Roth: Kövületek
Kivonat A. Roth: Gyökereink c. könyvéből 185-192 oldal
Ariel A. Roth biológus, a zoológia doktora.
Amit a kövületek elmondanak az evolúcióról
„Elfelejtünk egészen az alapokig ásni. Nem tesszük a kérdőjelet eléggé mélyre.”
(Ludwig Wittgenstein1)
A kövületek sok mindenről árulkodnak a Szentírás kontra tudomány régóta fennálló kérdésében. „Legfelsőbb bíróságnak kiáltották ki őket, ahol is az evolúció tana a vádlottak padjára kerül.”2 Mit is kell ezeknek a kövületeknek valójában elmondani az evolúcióról? Valóban olyan jól támogatják az evolúciót? Két fontos kérdést fogunk megvizsgálni,
– az evolúciós változások sebességét,
– és a kövületcsoportok kapcsolatát.
Az evolúciós változások sebessége és a kövületemlékek

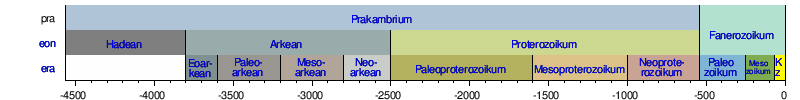
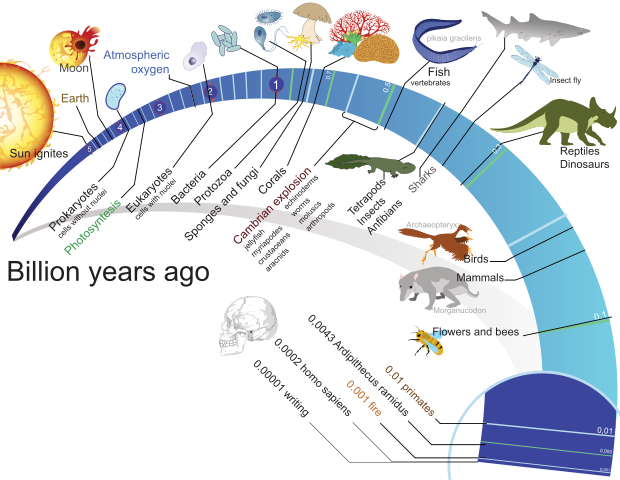
Néhány fontos kövületlelet, mint például a prekambrium legnagyobb részének nagyon egyszerű formái, amik az összetett állati kövületek alatt helyezkednek el, ideértve az Educara- és Burgess-pala3 szervezeteit, kétségbe vonja az általános evolúciós folyamat szokásos feltevését. Először is, mint arányaiban igen szabálytalan sebességű működést kellene megfigyelnünk az evolúciót.
Az evolúciós modell szerint az élet legalább 3500 millió évvel ezelőtt fejlődött ki, de közel 3000 millió évig viszonylag egyszerű egysejtű formában maradt. Aztán hirtelen, mintegy 100 millió év alatt, az állatvilág csaknem minden (körülbelül 40) törzse4 kialakult az úgynevezett kambriumi robbanás következtében, és gyakorlatilag egyetlen állattörzs sem alakult ki ezután. A 100 millió év időtartam a kambriumi robbanásra evolúciós szempontból egy bő intervallum. Egyesek csak 5-10 millió évet mondanak az állattörzsek többségének kialakulására, ami kevesebb, mint az evolúció teljes, feltételezett időtartamának (1/300) része. A Massachusettsi Műszaki Egyetemről Samuel Bowring ezt mondja: „ Amit szeretnék megkérdezni biológus barátaimtól, az az, hogy milyen gyorsaságot érhet el az evolúció, mielőtt kényelmetlenül kezdenék érezni magukat.”5 A kutatók a kambriumi régió moszatainak jelentős szaporulatáról is jelentést tesznek.6 Általában a növények, amik csak az egynegyedét képviselik a ma élő fajoknak, magasabban jelennek meg, különböző csoportokkal, a különböző szinteken. Az oszlop felsőbb részében folytatódik a maradványok hirtelen megjelenése. Például az emlősrendek legtöbbje feltehetően csupán 12 millió évvel ezelőtt jelent meg (az alsó harmadidőszakban). Az evolucionista Steven M. Stanley kihangsúlyozza, hogy mivel az átlagos történelmi időszakok előtti emlős fajok több mint egymillió évig fennmaradtak, csak 10-15 egymást követő fajnemzedéknek (chronospeciesnek) jutott ideje olyan eltérő emlőssé kifejlődnie, mint a bálna és a denevér. Azt mondja: „Ez teljességgel lehetetlen.”7, és olyan alternatívákat közöl, mint például a gyors változások s szabályozógénekben, és kis populációk, amelyekben a mutációk gyorsabban felszínre kerültek, elősegítve annak megértését, hogyan tudott az emlőstípusok nagy változatossága hirtelen, ilyen rövid idő alatt kialakulni. Egy „különleges, robbanásszerű evolúciót” jelentettek a madarakra vonatkozóan is, minden élő rendjükkel együtt, amelyek „mintegy 5-10 millió év” alatt alakultak ki.8 Mát korábban jeleztük, hogy a meg-megszakított egyensúly modellje nem megoldás.9 Az a fajok szintjén foglalkozik a változásokkal, és nem nevezi meg a nagyobb csoportok (rendek, osztályok, állat- és növénytörzsek) gyors eredetének problémáját.
Valójában a kövületemlékek egyszerűsödést mutatnak mind az állatok, mind a növények alaptípusai szempontjából az alsó fanerozoikumi szakasztól kezdve. Stephen J. Gould kiemeli, hogy jelentősen több alapállatfajta létezett a kambriumi rétegekben, mint mostanság. Kijelenti, hogy a hagyományos evolúciós törzsfamotívumot, ami egy egyszerű eredeti típussal (a törzzsel) kezdődik és egyre többféle szervezettel (az ágakkal és levelekkel) folytatódik, meg kellene fordítani, mivel kevesebb anatómiai formát találunk most, mint a múltban.10
A paleobotanikus Wilson Stewart és Gar Rothwell 31 „fő növénycsoportot” sorol fel az alsó paleozotikumi rétegekből, a ma létező, csupán 23-hoz képest.11
 A fő szervezetek területén is nagyobb változatosságot látunk az alsó paleozoikumi szakaszban, ahol 67 csoport jelenik meg a paleozoikumi részben, és csak 42 fentebb, a kainozoikumi részben. Ez a különbség még nagyobb lehet, mert a szám nem szól a paleozoikumi szakasz több kisebb növénycsoportjáról. A magasabban fekvő rétegekben a rétegtani oszlopban több faj is lehet12, de ez csak az alaptípusok kisebb változatait foglalja magában. Más szóval, több alapmotívum jelenik meg az alsó oszlop részében, de kevesebb motívum több változata uralkodik a felsőbb régiókban. A kipusztulások miatt kevesebb alapvető anatómiai forma maradt fenn az oszlop magasabban fekvő részében, pedig azt várnánk, hogy az evolúció fokozatosan többet hozott létre, ahogy az idő haladt.
A fő szervezetek területén is nagyobb változatosságot látunk az alsó paleozoikumi szakaszban, ahol 67 csoport jelenik meg a paleozoikumi részben, és csak 42 fentebb, a kainozoikumi részben. Ez a különbség még nagyobb lehet, mert a szám nem szól a paleozoikumi szakasz több kisebb növénycsoportjáról. A magasabban fekvő rétegekben a rétegtani oszlopban több faj is lehet12, de ez csak az alaptípusok kisebb változatait foglalja magában. Más szóval, több alapmotívum jelenik meg az alsó oszlop részében, de kevesebb motívum több változata uralkodik a felsőbb régiókban. A kipusztulások miatt kevesebb alapvető anatómiai forma maradt fenn az oszlop magasabban fekvő részében, pedig azt várnánk, hogy az evolúció fokozatosan többet hozott létre, ahogy az idő haladt.
 Egy ilyen szabálytalan evolúciós változási sebesség azt jelenti, hogy ha a változások megjelentek, minden bizonnyal gyorsan mentek végbe. A kövületek a kambriumi rétegek alatt kisebb evolúciós aktivitást jeleznek a geológiai idő első öthatod része alatt. Az ezt követő fejlődés bizonyára szakaszos sémát követet volna, magába foglalva a meg-megszakított egyensúlyt, gyakori nyugalmi időszakokkal a gyors evolúciós változások időszakai között. Ez viszonylag kevés időt hagyna az evolúciós változások tényleges folyamataira; néhány evolúciós modell szerint valószínűleg kevesebbet, mint a geológiai időskála 1 százalékát. Ilyen minták a kövületleletekben jelentősen csökkentik az évmilliók ezreit, amit a teljes evolúciós folyamatra megállapítottak. Az idő hiánya miatt ezek a szempontok növelik a komoly valószínűség eshetőségét, amivel az evolúció szembenéz.13
Egy ilyen szabálytalan evolúciós változási sebesség azt jelenti, hogy ha a változások megjelentek, minden bizonnyal gyorsan mentek végbe. A kövületek a kambriumi rétegek alatt kisebb evolúciós aktivitást jeleznek a geológiai idő első öthatod része alatt. Az ezt követő fejlődés bizonyára szakaszos sémát követet volna, magába foglalva a meg-megszakított egyensúlyt, gyakori nyugalmi időszakokkal a gyors evolúciós változások időszakai között. Ez viszonylag kevés időt hagyna az evolúciós változások tényleges folyamataira; néhány evolúciós modell szerint valószínűleg kevesebbet, mint a geológiai időskála 1 százalékát. Ilyen minták a kövületleletekben jelentősen csökkentik az évmilliók ezreit, amit a teljes evolúciós folyamatra megállapítottak. Az idő hiánya miatt ezek a szempontok növelik a komoly valószínűség eshetőségét, amivel az evolúció szembenéz.13
Míg a kövületleletekből a nagyobb evolúciós változások gyors megjelenését tételezzük fel, a kövületek egyéb adatai arra utalnak, hogy az evolúciós változás igen lassú lehetett. Néhány élő szervezet meglehetősen hasonlít a kövületmásához. Alsó devonkori atkák, amikről azt feltételezték, hogy 400 millió évvel ezelőtt fejlődtek ki, nagyon hasonlóak a mai fajokhoz.14 J.Willim Schopf több kékmoszat- (cianobaktérium-)kövületre talált rá a Bitter Springs sziklában, Közép-Ausztráliában, ami feltételezetten 850 millió éves, és ami azonosnak tűnik a ma élő fajokkal. Jelentést tesz még mintegy 90, különböző korúnak vélt ősi fajról, amelynek hasonmásai napjainkban is élnek.15 Wilson Stewart és Gar Rothwell hasonló szervezetekről számol be a késő archaikum és a középső proterozoikum közötti időből (1200-1700 millió évvel ezelőttről); ezt jegyzik meg: „Bár kevés határozható meg biológiai rendszereik evolúciójának sebességét illetően, nyilvánvaló, hogy morfotípusuk (alakjuk) meglehetősen változatlan marad a prekambriumon keresztül napjainkig.”16
Néhány életforma a Gunflint-kova korából az észak-amerikai Nagy –tavak vidékéről, ahol a rétegek kifejezetten közel 2000 millió évesek, igen hasonló a ma élő másához. Általánosan szólva, Andrew Knoll ezt állapítja meg: „Sok késő proterozoikumi prokarióta (sejtmag nélküli) morfológia, fejlődés és viselkedés szempontjából csak kicsit tér el az élő cianobaktérium-populációktól.”17
Az evolucionisták megpróbálják a változások hiányát vagy egy epizodikus (szabálytalan) evolúciós változási sebesség alapján megmagyarázni, vagy belső evolúciós változásokkal, amik nem láthatók, de a teremtés elméletén belül ezek a hasonlóságok a ma élő szervezetek kőzetekbe való beszivárgásának is lehetnek eredményei.18
Az a tény, miszerint ma az ev olúció elmélete az evolúció sebességét az extrém lassútól az extrém gyorsig egyaránt elfogadja, hogy megfeleljen a kövületeknek, jól illusztrálja azt, hogyan tud az általános evolúcióelmélet könnyen alkalmazkodni a sokféle adathoz. Az igen változó evolúciós sebesség kétségbe vonja a lassú, fokozatos evolúciós folyamat hagyományos nézetét, és csodálkozhatunk, miért van az, hogy néhány baktérium vagy hasonló egyszerű szervezet egészen emberré alakul 600 millió év alatt, míg más szervezetek 2000 millió éven keresztül a változás jelét sem mutatják.
olúció elmélete az evolúció sebességét az extrém lassútól az extrém gyorsig egyaránt elfogadja, hogy megfeleljen a kövületeknek, jól illusztrálja azt, hogyan tud az általános evolúcióelmélet könnyen alkalmazkodni a sokféle adathoz. Az igen változó evolúciós sebesség kétségbe vonja a lassú, fokozatos evolúciós folyamat hagyományos nézetét, és csodálkozhatunk, miért van az, hogy néhány baktérium vagy hasonló egyszerű szervezet egészen emberré alakul 600 millió év alatt, míg más szervezetek 2000 millió éven keresztül a változás jelét sem mutatják.
A kövületek a legjobb evolúciós modellnél is felfedik az igen szabálytalan evolúciós sebesség arányát. A lassú vagy evolúció nélküli elnyújtott periódusok, amiket a kövületek jeleznek, kevés időt hagynak a geológiai múltban igen valószínűtlen, összetett evolúciós változásokra.
A kövületleletek közötti hiányzó részek
 Amikor végzős hallgató voltam az egyetemen, az evolúcióprofesszor tájékoztatott engem, hogy a zoológiai tanszék testülete érdeklődik kreacionista hitem felől. Megkérdezte, hogy kifejteném-e nekik. Azt feleltem, arra figyeltem fel, hogy miképpen vezet egy bizonyos gondolatmenet az evolúcióban való hitre, de van néhány kérdésem a teóriát illetően. Ez felkeltette az érdeklődését. Az egyik érv, amit előhoztam, az volt, hogy nem értem, milyen úton fejlődhetett ki a teknős más hüllőkből anélkül, hogy fosszilis formát hagyott volna maga után. A teknős egy páratlan szervezet, és ennek a páratlanságnak a kifejlődésében — különösen a páncélnál — sok közbülső állomásnak kellene lennie, és mégsincs nyoma ennek a kövületekben. A paleontológusok teknőskövületek ezreit találták meg, volt közöttük négy méter hosszú is. Feltehetően több mint 20 millió évvel ezelőtt fejlődtek ki, de az alsó rétegekben, ahol először megjelennek, nem látjuk különleges páncéljuk kifejlődésének fokozatait.19 Miután megvitattunk más elgondolásokat is, a professzor elégedettnek tűnt a válaszaimmal, és egyetértett azzal, hogy az evolúcióelmélet hagy kívánnivalót maga után. Később megtudtam, az egyetlen ok, hogy a tantestület engedélyezte lediplomázásomat az volt, hogy nem tudtak megegyezni, mit csináljanak velem!
Amikor végzős hallgató voltam az egyetemen, az evolúcióprofesszor tájékoztatott engem, hogy a zoológiai tanszék testülete érdeklődik kreacionista hitem felől. Megkérdezte, hogy kifejteném-e nekik. Azt feleltem, arra figyeltem fel, hogy miképpen vezet egy bizonyos gondolatmenet az evolúcióban való hitre, de van néhány kérdésem a teóriát illetően. Ez felkeltette az érdeklődését. Az egyik érv, amit előhoztam, az volt, hogy nem értem, milyen úton fejlődhetett ki a teknős más hüllőkből anélkül, hogy fosszilis formát hagyott volna maga után. A teknős egy páratlan szervezet, és ennek a páratlanságnak a kifejlődésében — különösen a páncélnál — sok közbülső állomásnak kellene lennie, és mégsincs nyoma ennek a kövületekben. A paleontológusok teknőskövületek ezreit találták meg, volt közöttük négy méter hosszú is. Feltehetően több mint 20 millió évvel ezelőtt fejlődtek ki, de az alsó rétegekben, ahol először megjelennek, nem látjuk különleges páncéljuk kifejlődésének fokozatait.19 Miután megvitattunk más elgondolásokat is, a professzor elégedettnek tűnt a válaszaimmal, és egyetértett azzal, hogy az evolúcióelmélet hagy kívánnivalót maga után. Később megtudtam, az egyetlen ok, hogy a tantestület engedélyezte lediplomázásomat az volt, hogy nem tudtak megegyezni, mit csináljanak velem!
Olyan kérdések, mint a teknős eredete, százszor is megismétlődhetnek. A rétegtani oszlop minden, egymást követő részében fellelhetjük a sok új szervezetfajta hirtelen megjelenését. Az ezek ősei utáni kutatás az alsó rétegekben szinte teljesen eredménytelen. Charles Darwin teljesen tudatában volt ennek a problémának. A fajok eredetében azt mondja: „De amilyen mértékben ez a kihalási folyamat hatalmas méreteket öltött, a közbenső változatok számának is, amik korábban léteztek, épp oly óriásinak kell lennie. De akkor miért nincs minden geológiai képződmény és minden réteg tele ilyen közbeeső láncszemmel? A geológia biztosan nem fed fel semmiféle ilyen nagyszerűen fokozatos, szervezetekből álló láncolatot; és ez talán a legnyilvánvalóbb és legkomolyabb kifogás, ami felhozható a teória ellen.”20 Darwin továbbmegy, hogy a problémát a geológiai adatok „túlzott hiányosságának” tulajdonítsa. Azonban, hogy ő maga is elismerte, korának több vezető szakértője, mint „Agassiz, Pictet, és mindenki mást megelőzve professzor Sedgwick”, szembe szállt nézetével.21
 A hiányzó átmeneti formák általános képe nem változott jelentősen Darwin ideje óta. 120 évvel később David M. Raup, a chicagói Field Természetrajzi Múzeum vezetője és a Paleontológiai Társaság volt elnöke, megfigyelte, hogy „az élet fokozatos feltárásának megtalálása helyett, amit a Darwin idejében élt geológusok és a mai geológusok találnak: egyetlen, szaggatott emlékanyag; azaz, a fajok nagyon hirtelen jelennek meg a kövületsorrendben, kevés vagy semmilyen változást nem mutatnak létezésük alatt a maradványban, aztán hirtelen eltűntek a leletből.”22
A hiányzó átmeneti formák általános képe nem változott jelentősen Darwin ideje óta. 120 évvel később David M. Raup, a chicagói Field Természetrajzi Múzeum vezetője és a Paleontológiai Társaság volt elnöke, megfigyelte, hogy „az élet fokozatos feltárásának megtalálása helyett, amit a Darwin idejében élt geológusok és a mai geológusok találnak: egyetlen, szaggatott emlékanyag; azaz, a fajok nagyon hirtelen jelennek meg a kövületsorrendben, kevés vagy semmilyen változást nem mutatnak létezésük alatt a maradványban, aztán hirtelen eltűntek a leletből.”22
Éppen néhány évvel ezelőtt a paleontológus David B. Kitts, az Oklahomai Egyetem munkatársa bevallotta: „A fényes ígéretek ellenére, miszerint a paleontológia ki tuja mutatni a »látható« evolúciót, az evolucionistáknak néhány komoly nehézsége támadt; ezek közül legközismertebb a »hézagok« jelenléte a kövületleletek között.” Az evolúció megkívánja a fajok közötti átmeneti formákat, a paleontológia viszont nem szolgál ilyenekkel.23
Stephen Jay Gould ugyanezt hangoztatja. „Az átmeneti formák extrém ritkasága a fosszilis leletanyagok között a paleontológia üzleti titkaként áll előttünk. Az evolúciós törzsfáknak, amik a tankönyveket ékesítik, csak az ágai végén és csomóin vannak adatok; a többi — akármilyen ésszerű is — csak következtetés, nem pedig a kövületek bizonyítéka”.24
A kövületsorrend mintái kényszerítették az evolucionistákat, hogy elismerjék, az evolúció gyors kitöréssel történik. Azt is sugallták, hogy a változások kisebb populációkban történtek, ahol az átmeneti formák kövületként történt megőrződésének esélye sokkal kisebb, tehát a meg-megszakított modellből indulnak ki.25 Ezek a kijelentések magyarázatul szolgálnak az átmeneti formák hiányára vonatkoztatva, a közeli kapcsolatban levő fajok között, viszont nem nevezik meg a szervezetek főbb csoportjai közötti átmenetek hiányának sokkal jelentősebb problémáját.
 Az élő és kövületszerkezetek nagyobb kategóriákba sorolhatók, amit állta- és növénytörzseknek nevezünk. Ezek a fő elkülönülő csoportok a hierarchikus osztályozási rendszerben. A több mint egymillió különböző élő faj kevesebb mint 80 főbb csoportba (állat- és növénytörzsbe) sorolható. Miért különülnek el a csoportok? És amikor megnézzük a kövületeket, miért nem találjuk meg a fejlődésben levő átmeneteket az elkülönülő csoportok között? Itt bukik el az evolúciós modell, a legsúlyosabb tesztjénél. A remény egy evolúciós csodára, ami majd átalakítja az egyik alapfajt egy másikba, bizonyítatlan marad.
Az élő és kövületszerkezetek nagyobb kategóriákba sorolhatók, amit állta- és növénytörzseknek nevezünk. Ezek a fő elkülönülő csoportok a hierarchikus osztályozási rendszerben. A több mint egymillió különböző élő faj kevesebb mint 80 főbb csoportba (állat- és növénytörzsbe) sorolható. Miért különülnek el a csoportok? És amikor megnézzük a kövületeket, miért nem találjuk meg a fejlődésben levő átmeneteket az elkülönülő csoportok között? Itt bukik el az evolúciós modell, a legsúlyosabb tesztjénél. A remény egy evolúciós csodára, ami majd átalakítja az egyik alapfajt egy másikba, bizonyítatlan marad.
A paleontológusok valószínűleg sok új kövületfajt találnak majd a jövőben, de ahogy az már évszázadok óta történik, számíthatunk rá, hogy valamely különálló főbb csoportokhoz fognak tartozni.26 Állíthatjuk, ahogy Darwin is tette, hogy a fosszilis leletanyag tökéletlen, de kövületek millióit gyűjtötték már be. Azt, hogy miért osztják fő csoportokra folyamatosan a kövületeket, amíg a közöttük levő nagy hézagok eltűnnek, az evolucionistáknak nehéz megmagyarázni. Nem tűnik lehetségesnek, hogy a katasztrófaesemények vagy balesetek — elősegítve a kövülés folyamatát és a megőrződést — csak akkor történnek meg, amikor éppen nem fordul elő fejlődés a fő csoportok között.
A tiszteletre méltó harvardi paleontológus, George Gaylord Simpson ismertette az átmeneti formák csökkenő számának problémáját az osztályozási rendszerben felfelé haladva. (…)27 az evolúciós modell szerint az átmeneti formák legnagyobb számát a fő csoportok között várhatnánk, éppen ott, ahol nevezetesen hiányoznak.
Néhány példa illusztrálja a hiányzó láncszemek problémáját.28
A kambriumi robbanás nemcsak egy esete az összes nagyobb állattörzs kialakulásának, ami közel azonos helyen jelenik meg a rétegtani oszlopban. Ez egy ősök nélküli helyzet is, ami elmondhatná, hogyan fejlődhettek ki. A paleontológusok alaposan tanulmányozták a kőzeteket közvetlenül a kambriumi robbanás alatt, amelyben azt várták, hogy megtalálják az átmeneti formákat. Ez gyakorlatilag eredménytelen keresés volt. A kövületbizonyítékok hiányában a paleontológusok dilemmával küszködnek annak tekintetében, hogy miképpen állhatnak a csoportok egymással rokonságban. Frederick Schram, a Scripps Oceanográfiai Intézet munkatársa ezt mondta: „Valószínűleg egyetlen témára sem ennyire jellemző az egyéni spekuláció, mint a gerinces állatok törzsének rokonsága. Talán nincs is két szakértő, akinek a véleménye megegyezne. Továbbá az egyéni aspektusok versengő értelmezéseinek bősége a gerinctelenek anatómiájáról, valamint a nevek zavaró sora, mindenféle „feltételezett” ősre és kitalált állatokra vonatkoztatva, félelmetes.”29
A növények evolúciójával kapcsolatos kérdések nem sokban különböznek. (…) Harold C. Bold (Texasi Egyetem) és szerzőtársai azt állították, hogy ők „az összehasonlító alaktan, sejttan, biokémia és kövületemlékek jelenleg rendelkezésre álló bizonyítékának gondos mérlegelése után, jelenleg nem hajlandók egybeolvasztani semelyik kettő vagy több növénytörzset abból a 19-ből, amelybe ők már próbaképpen besorolták a növényvilág szervezeteit”. 30
Például a virágos növények teljesen kialakulva és bőségükben jelennek meg hirtelen a kövületleletek között. Darwin a virágos növények eredetét „gyűlöletes rejtélynek” nevezte. Több mint egy évszázaddal később vezető paleontológusok (Axelrod, Bold, Knoll és Rothwell) még mindig „gyűlöletesnek” nevezték a problémát.
 A repülni tudó szervezeteket négy fő csoportra osztották: rovarok, pteroszauruszok (repülő hüllők), madarak és denevérek (bocsánatot kérek az emberiségtől és a légiipartól!). A repülés egy igen különös működési folyamat, amihez a szárnyak mellett sok más tulajdonság is kell. Például a kisrepülőgépek szerkezete határozottan különbözik az autókétól. Természetesen a repülés fokozatos fejlődésétől elvárjuk, hogy nyomot hagyjon a kövületemlékek között. De amikor a rovarkövületek először megjelentek a rétegtani oszlopban, a repülés már teljesen kifejlődött.32
A repülni tudó szervezeteket négy fő csoportra osztották: rovarok, pteroszauruszok (repülő hüllők), madarak és denevérek (bocsánatot kérek az emberiségtől és a légiipartól!). A repülés egy igen különös működési folyamat, amihez a szárnyak mellett sok más tulajdonság is kell. Például a kisrepülőgépek szerkezete határozottan különbözik az autókétól. Természetesen a repülés fokozatos fejlődésétől elvárjuk, hogy nyomot hagyjon a kövületemlékek között. De amikor a rovarkövületek először megjelentek a rétegtani oszlopban, a repülés már teljesen kifejlődött.32
A repülő pteroszauruszok, madarak és denevérek tökéletesen működő repülő szervezetekként jelentek meg hirtelen. A repülés kifejlődéséhez szükséges anatómiai változások, ideértve a csontozatban, az izomzatban, tollazatban, légzésben és idegrendszerben való átalakulásokat, hosszú időt vennének igénybe, és az ilyen változásokon keresztülment szervezetek biztosan hátrahagynának a kövületekben valamiféle nyomot a közbenső fázisokból. A madarak tolla feltehetően az ősi hüllők pikkelyéből fejlődött ki. Bárki, aki mikroszkóp alatt megfigyelte a madártollat, felismeri, hogy az igen bonyolult és különleges szerkezet. Vajon a részek hüllőpikkelyből történt kialakulásának kiterjedt folyamata egy irányítatlan evolúció által, ami sikeres fejlődési vonalat is tartalmaz, nem hagyott volna nyomot a kövületekben? Ez idáig azonban egy ilyet sem találtak.
Hivatkozások
1. Wittgenstein L. 1980. Culture and value. Ford.: Wright G. — Chicago University of Chicago press 62e o. Eredeti címe: Vermischte Bemerkungen
2. Lull R.S. 1935. Fossils: what they tell us of plants and animals of the past, 2.kiad. New York University Society 3.o.
3. Lásd 9. fej.
4. Az optimista paleontológusok azt mondták, hogy a kambriumi robbanásnál már 100 állattörzs létezett. Lásd: Lewin R. 1988. A lopsided look at evolution. Science 241_ 291-293.
5. a) Bowring S. A. Grotzinger J. P., Isachasen C. E. Knoll A. H. Pelechaty S. M., Kolosov P. 1993. Calibrating rates of Early Cambrian evolution, Science 261: 1293-1298. Az idézet forrása: b) Nash M. 1995. When life exploded. Time 146 (23):66-74.
6. Kerr R.A. 1995. Timing evolution’s carly bursts. Science 267: 33,34.
7. Stanley S. M. 1981. The new evolutionary timetable: fossils, genes, and the origin of the species. Nem York: Basic Books, 93.o.
8. Feduccia A. 1995. Explosive evolution in Tertiary birds and mammals. Science 267.637.638.
9. Lásd a 8 fej.
10. a) Gould S. J. 1989. Wonderful life: The Burgess Shale and the nature of history. New York and London: W.W. Norton and Co., 39-50.o. Ahogy várható is volt, az elmélet nem maradhatott kritika nélkül. Lásd b) Briggs D. E., Fortey R. A., Wills M. A. 1992. Morphological disparity int he Cambrian. Science 256: 1670-1673. ; későbbi megvitatása: c) Foote M., Gould S. J. and Lee M. S. Y. 1992, Cambrian and recent morphological dispary, Science 256: 1816-1817, with response by Briggs, Fortey, and Wills in Science 256: 1817,1818.
11. Stewart W. N., Rorhwell D.W 1993. Paleobotany and the evolution of plants. 2. kiad. Cambridge and New York: Cambridge University Press, 510-511.o.
12. Volt olyan nézet, hogy faji sokféleség a gerinctelenek között igencsak összefüggésben áll az üledékes kőzetek területével és mennyiségével. Lásd: a) Raup 1976. Species diversity in the Phanerozoic: an interpretation. Paleobiology 2: 289-297 – b) Raup 1972. Taxonomic diversity during the phanerozoic, Science 177: 1065-1071. Ugyanis az üledékek mennyisége és feltártsága nagyobb a geológiai oszlop magasabban fekvő rétegeiben, és ez a következtetéséket a magasabban fekvő oszloprészben feljegyzett fajok nagyobb száma irányában befolyásolja. Az alaptípusokból kevesebbet fedeztek fel.
13. Lásd: 4-8 fej.
14. Bernini F. 1991. Fossil Acarida, Simonetta A. M. Conway Morris S. szerkesztők. Cambridge and Vew York ; Cambridge University Press, 253-262.o.
15. a) Pennisi E. 1994. Static evolution: is pond scum the same now as billions of years ago? Science News 145: 168,169; b) Schopf J. W. 1968. Microflora of the Bitter Springs Formation, Late Precambrian, central Australia. Journal of Paleontology 42: 651-688.
16. Stewart and Rothwell, 44.o. (11. megj.)
17. Knoll A.H. 1990. precambrian evolution of prokaryotes and protists. Briggs D:E. G., Crowther P.R. szerkesztő. Paleobiology: a synthesis. Oxford and London: Blackwell Scientific Publications, 9-16.o.
18. Lásd a 10.fej.
19. a) Caroll R. L. 1988. Vertebrate paleontology and evolution. New York: W. H. Freeman and Co. 207. o. A teknős evolúciójának megmagyarázására tett kísérletért embriológiai alapokon és nem paleontológiai adatok alapján lásd b) Petto A. J. 1983. The turtle: evolutionary dilemma or creationist shell game? Creation/Evolution 3 (4): 20-29. a csontokon alapuló anatómiai magyarázat kísérletére lásd: c) Lee M. S. Y. 1993. The origin of the turtle body plan: bridging a famous morphological gap. Science 261: 1716-1720
20. Darwin C. 1859. The origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. London: John Murray. Burrow J. W. szerkesztő. 1968. reprint. London and New York: Penguin Books, 291-292.o.
21. Uo. 309.o.
22. Raup D. M. 1979. Conflicts between Darwin and paleontology. Field Museum os Natural History Bulletin 50: 22-29.
23. Kitts D. B. 1974. Paleontology and evolutionary theory. Evolution 28: 458.
24. Gould S. J. 1980. The panda’s thumb: more reflections in natural history. New York and London. W.W. Norotn and Co. 181.o.
25. Lásd 8. fejezet.
26. Cowen gondolata az, hogy jó csontozatú sekély tengeri állatok összes törzsét felfedezték. Cowen R. 1995. History of life. 2. kiad. Boston, Oxford, and London: Blackwell Scientific Publications, 97.o.
27. Simpson G.G. 1967. The meaning of evolution: a study of the history of life and of its significance for man. Jav. kiad. New Haven and London: Yale University Press, 232-233.o.
28. Az evolucionisták és a kreacionisták és mások is sokat írtak ezekről a hézagokról. Néhány példa, amely felismeri a problémát: a) Denton M. 1985. Evolution: a theory in crisis. London: Burnett Books; b) Grassé P.P. 1977. Evolution of living organisms: evidence for a new theory of transformation. Carlson B. M. Castro R. fordítók. New York, San Francisco, and London: Academic Press. Eredeti cím: L’Évolution du Vivant… c) Hitching F. 1982. The neck of the giraffe: where Darwin went wrong. New Haven and New York: Ticknor and Fields; d) Oxford: Oxford University Press: e) Johnson P.E. 1993. Darwin on trial. 2. kiad. Downers Grove, III. InterVarsity Press; f) Lovtrup S. 1987. Darwinism: the refutation of myth. London, New York, and Sidney: Croom Helm; g) Pitman M. 1984. Adam and evolution. London, Melbourne, and Sidney: Rider and Co.
29. Schram F. R. 1991. Cladistic analysis of metezoan phyla and the placement of fossil problematica. Simonette and Conway Morris, 35-46.o. (14. megj.)
30. Bold H. C. Alexopoulos C. J. Delevoryas T. 1987. Morphology of plants and fungi. 5. kiad. New York and Cambridge: Harper and Row, 823.o.
31. a) Axelrod D. I. 1960. The evolution of flowering plants. Tax S. szerkesztő. The evolution of life: its origin history and future. Evolution after Darwin: The University of Chicago centennial, 1k. Chicago: University of Chicago Press, 227-305.o., b) Bold H. C. 1973. Morphology of plants, 3. kiad., New York and London. Harper and Row, 601.o. ( a 4. és 5. kiadását ennek a szövegnek még két társszerző alkotta, és a „visszataszító” szót már nem használták, bár az elmélet még mindig uralkodó a könyvben.; c) Knoll A. H. Rothwell G. W. 1981. Paleobotany: perspectives in 1980. Paleobiology 7 (1) 7-35.o.
32. Wootton R. J. 1990. Flight: arthropods. Briggs and Crowther, 72-75.o. (17. megj.)
A könyv részleteit az Advent Kiadó engedélyével és szíves közreműködésével tesszük közzé. Az Advent Kiadó is fontos együttműködő partnerünk. A könyvet 1980 Ft-os áron az alábbi helyen lehet megrendelni: http://www.adventkiado.hu/
Bővebben: