EAH 2. Genetika és DNS
Részlet az Evolúció Akhilleusz sarkai (halálos pontjai) című könyvből

2. fejezet:
Genetika
és DNS
48-77 oldal
A könyvet Robert Carter, Ph.d szerkesztette.
Kiadó: Creation Book Publishers. Második kiadás, 2014. október
Honlap: Creation.com
Fordította: Dr. Cserháti Mátyás biológus.
Lektorálta: Dr. Hecker Kristóf biológus.
Dr Don Batten Ph.D: Az evolúció Akhilleusz sarkai című könyvének fejezetenkénti fordítása.
Tartalomjegyzék:
1. fejezet: Természetes szelekció
6. fejezet: Radiometrikus kormeghatározás
8. fejezet: Etika és erkölcsiség
2. fejezet

Dr. Robert Carter
Tengerbiológia doktora, Miami Egyetem
Dr. Carter, aki a korallzátonyok szakértője, a PhD kutatásának jó részét a medúzákban és korallokban lévő egyik fehérjecsaládnak – a zöld fluoreszkáló fehérjének (angol rövidítéssel GFP) és rokonainak – szentelte. A figyelme azóta az emberi genetika felé fordult. Az első munkája a legkorábbi emberi mitokondriális genom – gyakorlatilag Éva – konszenzus szekvenciájának meghatározása volt. Jelenleg az emberi genetika további vonatkozásait vizsgálja, beleértve a diverzitás és a mutáció mintázatait, és hogy azok hogyan kapcsolódnak a Mózes első könyvében bemutatott genetikai modellhez.
Miután fiatal korában többféleképpen próbálta a teremtéstörténetet az evolúcióval összeegyeztetni, elsőéves egyetemistaként végül elfogadta, hogy az evolúció „tény”. Később azonban rákényszerült, hogy újragondolja az álláspontját, így az evolúciót elvetette a bibliai teremtés kedvéért, de csak a doktori képzés első évében győződött meg teljesen a fiatal föld elmélet helyességéről.
Mint mély genetikai ismeretekkel rendelkező teremtéshívő, és mint volt evolucionista, még ha csak rövid ideig is, természetes választásnak tűnt felkérni, hogy a darwini evolúció második Achilles-ináról, a genetikáról írjon.
Lásd: creation.com/dr-robert-carter
Az első fejezetben láthattuk, hogy a természetes szelekció eredetileg nem Darwin ötlete volt. Azt is láttuk, hogy Darwin nagyszerű ötletét nem elég magyarázni azzal, hogy a közös eredet révén minden faj rokona egymásnak. Ebben a fejezetben olyasvalamit fogunk megvizsgálni, amiről Darwinnak fogalma sem volt – ez a genetika. Az ismeretének hiánya ellenére nem tudunk a természetes szelekcióra épülő evolúcióról az örökítő anyagban történő változások érintése nélkül beszélni. Az evolúció gyenge pontjainak megértésében kulcsfontosságúak azok a kérdések, mint „Hogyan adódnak át a gének az egyik generációtól a másikra?”, illetve „Mekkora mutációs rátát lehet egy populációban az idők során fenntartani?”
A genetika rövid története
A filozófusok évezredekig vitatkoztak arról, hogy az élőlények hogyan adják tovább utódaiknak a tulajdonságaikat. Az emberi történelem nagy részében nem tudtuk, hogy mi okozza a fajbeli változatosságot, sem azt, hogy ez a variáció hogyan adódik át két generáció között. A 19. század végéig a legtöbb ember a kevert öröklődésben hitt. Ez azt jelenti, hogy az utód a két szülő kombinációja vagy átlaga volt. Ez bizonyos értelemben kézenfekvő volt, mivel könnyű mindkét szülő tulajdonságát meglátni a gyerekekben, de ugyanakkor az is kérdés volt, hogy minden tulajdonság átlagos értéket vesz-e fel az idők során? Az emberek tudtak erről a problémáról, de jobb ötleteik nem voltak.
Mivel Charles Darwin nem a megfelelő korban élt, ezért nem tudott a genetikáról. Így az evolúcióról szóló elgondolásai egy olyan vákuumban keletkeztek, ahol sok spekuláció volt. Mivel nem volt megelégedve kora uralkodó elképzeléseivel, Darwin létrehozta a saját elméletét, amit pángenezisnek1 nevezett. Bizonyos „testecskéket” képzelt el, amit különböző testi szervek a környezeti ingerek hatására hoznak létre. Ezek a testecskék ezek után az ivarmirigyekbe vándoroltak, amíg tovább nem adódtak a következő generációba. Azonban sem fizikai, sem kísérletes bizonyíték nem állt rendelkezésre elmélete alátámasztására. Ugyanakkor egy régebbi elméleten alapult, amit lamarckizmusnak vagy lamarcki öröklődésnek neveztek. Jean-Baptiste Lamarck (1744-1829) azt tanította, hogy az egyed a környezeti hatásokra úgy válaszol, hogy erősebb, magasabb, távollátóbb lesz, vagy jobban ellenáll a túlzott hőségnek. Az egyed ezek után az újonnan megszerzett tulajdonságait az utódainak is továbbadja, amelyek aztán születésüktől fogva erősebbek vagy magasabbak lesznek, mint a szülők. Az ehhez a gondolathoz kötődő kifejezés „a szerzett tulajdonságok öröklődése”.
Darwin bizonyára hitt is ebben2, de tévedett. Darwin pángenezis elmélete azért is furcsa, mivel rögtön azután tette közzé (1868), hogy Gregor Mendel, a modern genetika atyja, felfedezte a genetikai öröklődés törvényeit (először 1866-ban közölte3). A kolostorának kertjében egyedül dolgozva Mendel azt találta, hogy sok tulajdonság diszkrét egységekben van jelen (fehér vagy lila virágok, sárga vagy zöld magvak, stb.), és minden variánst allélnak nevezett. Azt is megállapította, hogy minden egyed két allélt hord minden tulajdonsághoz (pl. fehér és fehér, lila és fehér, vagy lila és lila), és hogy mindkét szülő csak egy példányt ad tovább az utódának. Az utódok így nem a szülők átlagértékét képviselik (keveredés), hanem a szülők gyermekeiknek átadott diszkrét tulajdonságainak kombinációit jelenítik meg.
 Mendel ugyan szerzetes volt, de nem volt ismeretlen előtte a tudomány, mivel őt a fizikában és a filozófiában is egyetemi szinten képezték. Az ő korában az egyház egy olyan intézmény volt, ahol hagyományosan számos tudományágat ápoltak, amiről Darwin Cambridge-ben4 folytatott teológiai és tudományos képzése is tanúságot tesz, amikor még vidéki lelkésznek készült.
Mendel ugyan szerzetes volt, de nem volt ismeretlen előtte a tudomány, mivel őt a fizikában és a filozófiában is egyetemi szinten képezték. Az ő korában az egyház egy olyan intézmény volt, ahol hagyományosan számos tudományágat ápoltak, amiről Darwin Cambridge-ben4 folytatott teológiai és tudományos képzése is tanúságot tesz, amikor még vidéki lelkésznek készült.
Darwin ugyanazokkal az eszközökkel rendelkezett, mint Mendel, több ideje is volt, és több pénzt is költhetett a tudományos munkára, de nem dolgozott ki semmi olyasmit, ami a modern genetika elméletét megközelítette volna. Darwin részletesen leírta a sokféle növény- és állatfajban lévő genetikai variációs mintázatokat, és még Mendel híres 3:1-es vad:recesszív típusok arányát5 is megfigyelte egy alkalommal, kettős hibrid keresztezés során.6 Darwin több esszét és cikket írt ezekről a témákról.7 De akkor miért nem fedezte fel a genetikát? Talán sohasem lehetünk ebben biztosak, de Darwin azt szerette volna, ha a fajok örökké változatosak lennének, mivel előzetesen már a fajok folyamatos eloszlása mellett tört pálcát, ahol számtalan kapcsolatot talált a fajok között, visszanyúlva a végtelenségig. Ez szükségszerűen a tulajdonságok folyamatos eloszlását követelte meg, mivel a pángenezis elmélete a lamarckizmuson alapszik. De a gének nem a folyamatos eloszlást követik. Diszkrét tulajdonságokat kódolnak. Noha igaz, hogy sok tulajdonság sok különböző gén additív hatása révén jön létre (ami folyamatos eloszlásnak tűnhet), azonban ez a környezet és a genetika közti kapcsolatot még bonyolultabbá teszi. Darwin nem jó irányban keresgélt. Mendel kísérleti munkát végzett, és ő a helyes megoldást találta meg. Darwin elméleti szakember volt, és pusztán filozófiai okok miatt tévedhetett.8
Azonban ez nem csak Darwin hibája volt, mivel abban az időben sokan nem látták ezt. Több évtized telt el, mire Mendel gondolatai ismertebbek lettek. Darwin viszont egész életében nem ismerte azokat az alapvető folyamatokat, amikre az evolúció elméletét kellett volna alapoznia. Kiderült ugyanis, hogy a genetika törvényei nem kedveznek az evolúció elméletnek.
A modern genetika
A 20. század fordulóját követően a genetika tanulmányozása egyre gyorsult. Felfedezték, hogy teljesen új tulajdonságok, ún. mutációk, jelenhetnek meg hirtelen egy egyedben, és ezzel Mendel munkásságát újra felfedezték. Sok időbe telt, de a húszas évekre a mutáció, a természetes szelekció és a genetika összevonva a modern szintézis elméletét teremtette meg. A keveredő öröklődést, a pángenezist, a lamarckizmust mind elvetették egy újabb felfogás kedvéért, amit úgy 150 évvel később sem lehet nagyon megkülönböztetni attól, amit a legtöbb evolucionista ma hisz.
A genetika tudománya a század első felében lassan , majd az ötvenes évektől robbanásszerűen fejlődött. 1952-ben a híres Hershey-Chase kísérlet azt bizonyította, hogy a DNS az öröklődés hordozója (erre volt bizonyíték előtte, de sok ember abban hitt, hogy ezt a fehérjék végzik). 1953-ban Watson és Crick felfedezte, hogy a DNS kettős spirál alakot vesz fel. A Meselson-Stahl kísérlet 1958-ban azt mutatta, hogy a DNS szálak elválnak egymástól és külön másolódnak. Nem sokkal később pedig a fehérje kódot is kidolgozták, és azt látták, hogy három DNS betű kódol egy aminosavat a fehérjében.
A modern genetika alapjait ezzel lerakták. A génszekvenálást a következő évtizedekben fejlesztették ki. Ezt aztán automatizálni tudták. Az ezredforduló pedig egy új világot hozott, mivel 2003-ban a Humán Genom Projekt az emberi sejtmagban lévő lineáris DNS 93%-át leírta. A merész cél és a hatalmas finanszírozási igény az amerikai adófizetőktől három milliárd dollárt követelt, de cserébe meghatározták a DNS három milliárd betűjét. Ez a nagyszerű (és költséges) eredmény sok mindent megváltoztatott. Ma a technológia gyors fejlődésével sok cég naponta könnyedén elő tudja állítani több humán genom megfelelőjét, egyenként csak néhány ezer dollárért, de néhány éven belül ezt is túlszárnyalják majd. A technológiai fejlődés lehetővé teszi az egyre gyorsuló felfedezéseket, és ezzel sok hagyományos elképzelés megkérdőjeleződik.
Az adatok gyorsan gyűlnek. Manapság olyan kérdésekre tudunk választ adni, amiről egy vagy két évtizeddel ezelőtt csak álmodtunk. Talán először a világtörténelem során, létrejött egy olyan tudományterület, ahol több az adat, mint az elmélet! Köszöntelek a modern genetika világában!
A DNS, mint információ tároló közeg
Minden élet fennmaradása és szaporodása a specifikus információtól (pontosabban egy információhalmaztól) függ. Az információ többségét a DNS kódolja, egy olyan molekula, ami párját ritkító információ tároló közeg. Rengeteg utasítást tudunk akár egy kis darab DNS-be beletenni. Gondoljunk egy nyomtatott oldalra. Az oldalon lévő minden egyes betű három vagy négy milliméter magas lehet, és több ezer betű van egy átlagos oldalon. Ha az emberi genomot szövegként kinyomtatnánk, és ha beletennénk Bibliaméretű könyvekbe (a fordítástól függően körülbelül 3,5 milliárd betű van egy átlagos angol nyelvű Bibliában), ehhez 850 Bibliára lenne szükség. És mivel egy példányt az anyádtól és egy másikat az apádtól kapod, két genomot tartalmaz a testedben lévő minden egyes sejt (a vörösvérsejtek kivételével, amelyeknek nincsen sejtmagjuk). A DNS betűi azonban csupán egy méter milliomod részét teszik ki. Ennél a méretnél mind a három milliárd betű könnyedén belefér olyasvalamibe, ami kisebb, mint ennek a mondatnak a végén lévő pont. A sejtben a DNS láncokat formál (kromoszómák), úgy, hogy ha sorba állítanánk, akkor kevesebb, mint 2 méter magas lenne. Ezek a hosszú, törékeny, ragadós szálak egy olyan sejtmagba tömörülnek össze, amely csak 6-10 μm (egy milliomod méter) átmérőjű. Egy ember felépítéséért és fenntartásáért szükséges információ egy olyan apró valamibe tömörül össze, amit csak mikroszkóppal lehet látni!
De ezek a szálak nem nagyon stabilak. Az egyszálú DNS nagyon törékeny. A sejtmagban minden szál egy másik szállal párosul, és ez a kettő egymás köré tekeredik úgy, hogy egy klasszikus kettős spirál alakot vesz fel. Egy DNS a valóságban két egymástól független, de egymást kiegészítő molekula, ami egymáshoz elektrosztatikusan kötődik. Az emberi genom három milliárd betűje 23 különálló kromoszómában található, amelyek 50 és 250 millió betű hosszúságúak. A DNS azonban még ebben a konfigurációban sem nagyon stabil. A DNS hihetetlenül érzékeny a degradációra, és azt becsülik, hogy egy átlagos testi sejtben naponta csaknem egymillió DNS lézió történik (sugárzás okozta szál törés, vagy egyes bázisok oxigén hatására történő megsemmisülése)9 – miközben 100 billió sejtünk van! Képzeld el azt a ráfordítást, amit egyetlen sejtnek minden nap el kell végeznie, hogy fenntartsa a DNS-ét. Sokszorozzuk meg ezt a mennyiséget! Mennyi az 1.000.000-szor 100.000.000.000.000? Ennyi kell ahhoz, hogy a tested a genomodat napi szinten fenntartsa.
A DNS törékenysége csak egy az evolúciós genetika sok Achilles-ina közül, de az egyik legfontosabb. Ahhoz, hogy a DNS hasznos legyen, a fenntartása rengeteg javítóenzimet igényel. A DNS sokféleképpen tud megsérülni, és minden sérülésre más és más speciális javító enzim van, de ami még nagyobb kihívást jelent az evolúciós modellre az az, hogy a DNS ezeket az enzimeket is kódolja, miközben a DNS-t nem lehet a sejtben nélkülük fenntartani. Ez a tyúk-vagy-tojás probléma tipikus példája! Ezek az enzimek nagyon érzékenyek a változásokra. A DNS javító illetve másoló enzimekben történő mutációk legtöbbször katasztrofálisak. Akkor hogyan is keletkezhettek mutációk és természetes szelekció révén az idők során? Nélkülük az élet nem tud létezni, de az élet nélkülük kellett hogy előálljon, a DNS-ben tárolva minden információt, mielőtt a DNS-eszköztár létrejött volna. Mivel az élet eredetéről a következő fejezetben fogunk bővebben szólni, most csak annyit árulunk el, hogy a DNS az utolsó molekula, amelyről azt gondolhatnánk, hogy az élet korai szakaszaiban információt tárolhatott.
A DNS és az információs elmélet
A DNS molekulában lévő nukleotidok sorrendje egy üzenetet alkot, tele információval.10,11 Egy üzenet továbbítható, de a valóságban nem lehet üzenetet feladó nélkül küldeni. Az élet rejtélye nem abban áll, hogy miért annyira komplexek azok a molekulák, amelytől az élet maga függ (noha ez egy nagy rejtély). Sőt, még csak nem is a részek komplex összeállítása, amit az élőlények használnak (bár ez is egy nagy rejtély). Nem, az élet legnagyobb rejtélye az, hogy hogyan keletkezett az információ, amelyre az élet épül. Amint az előző részben láttuk, ez már az élet eredetének kérdését érinti, ami nem ennek a fejezetnek a kérdése. Ami minket itt a leginkább érdekel az az, hogy hogyan tartjuk fent az információt a genomban az élőlény egész életén keresztül, illetve az egyik generációról a másikra.
Az információ fenntartása egy másik Achilles-ín az evolúciós genetika számára, mivel működésképtelen egy majdnem tökéletes információ-fenntartó rendszer nélkül. És még így is, a csodálatos DNS javítórendszerünkkel a testünkben is összeszedünk 100 új mutációt személyenként és generációnként.12 Úgy becsülik, hogy sejtosztódásonként 1-3 mutáció keletkezik.13 És ez nem is egy rossz hibaarány egy parányi molekuláris gépezetnél, ami a DNS gyors másolását végzi, mielőtt a sejt osztódna. De ha közben arra gondolunk, hogy hány sejtosztódásra van szükség ahhoz, hogy egy egész ember létrejöjjön, akkor arra jutunk, hogy minden ember minden lehetséges mutációt több példányban hordoz, és ennek a száma az életkorral tovább növekszik. A hibák felhalmozódása az idő során azt eredményezi, hogy egy hatvanéves embernek minden egyes bélsejtjében 40.000 mutáció van.14 Sőt, mi több, balesetek és betegségek nélkül is ez a rengeteg mutáció mindannyiunk halálához vezetne.
Szerencsére a Teremtő (Isten) csodálatos gépezetet hozott létre, hogy megvédje az emberiséget ezeknek a mutációknak a káros hatásától. Amikor a petesejt megtermékenyül, a zigóta korlátozott számú osztódáson megy keresztül, mielőtt a következő generáció sejtjei formálódnak. Egy leány csecsemőben pusztán 23 sejtosztódás történik, mielőtt a petefészek kialakul. A petefészekben lévő peték védve és táplálva vannak addig, ameddig a peteérés el nem kezdőik, akár 40 évvel később, miután többet nem osztódnak. A legtöbb ember nem is sejti, de amikor egy anya egy lánnyal várandós, három generáció van jelen egyszerre: az anya, a lánya, és a magzat petefészkében lévő petesejtek. A férfiaknál azonban más a helyzet. Egy fiú magzat körülbelül harminc sejtosztódáson megy keresztül az anyaméhben, mielőtt kialakulnak a szaporodás sejtjei, de a pubertás kortól ezek gyorsan osztódni kezdenek, és a halál bekövetkeztéig osztódnak. Ez azt jelenti, hogy egy apa több mutációt ad át egy olyan gyereknek, aki idős korában születik, mint egy olyannak, amely fiatal korában születik.15
Néhány, a szülőtől a gyerekig átadódó mutációkat vizsgáló tanulmány néhány meglepő eredményt mutatott fel. Kiderült, hogy azon mutációk száma, amely valamelyik szülőtől származik, változatos lehet, néha több származik az apától, néha több az anyától.16 Ez kihívást jelent az általános emberi genetikai történelem modelljeire nézve, amely egy olyan mutációs órát feltételez, ahol a mutációs ráta időben és az emberiség földrajzi eloszlását tekintve egyenlő. Ennek a következményeit a későbbiekben részletesebben fogjuk tárgyalni.
Noha e mutációk többsége nem katasztrofális (különben mindannyian már halottak lennénk), azért még károsak. A genetikusok ezeket kevéssé káros mutációknak hívják, és az emberi genomban történő felhalmozódásuk nagy kihívást jelent az evolúció elmélet számára.17 Ezek káros mutációk, és gyorsabban is adódnak hozzá az emberi genomhoz, minthogy a természetes szelekció elméletileg el tudná őket távolítani. Valóban, csakis a legkárosabb mutációkat „látja” a természetes szelekció (lásd első fejezetet). Így az evolúció visszafelé halad, és képtelen az élethez szükséges információ degradációját megfékezni. Ha ez igaz, akkor hogyan is keletkezhetett az információ? És a fajok hogyan maradhattak fenn évmilliókig anélkül, hogy elpusztultak volna?
A „hulladék” DNS (többnyire) működőképes!
Évtizedekig hallottuk a mesét. Valahogy így szól: „Az emberi genomnak csakis 2 vagy 3 százaléka funkcionális. A DNS maradék része haszontalan szemét, ami az evolúciós múltunkból maradt ránk.” Noha még sokan vallják ezt a nézetet, friss felfedezések kimutatták, hogy ez nem igaz. Akkor miért van az, hogy ezt olyan sokszor és annyi ideig hallottuk? Ez azért van, mert a biológiai evolúciónak szüksége van a hulladék DNS elméletre, hogy egy nagy matematikai problémát megoldjon.
 A késői ötvenes években a híres populációgenetikus, J. B. S. Haldane kimutatta, hogy a természetes szelekció nem tudja a jótékony mutációkat kiválogatni még az ember evolúciós történelme alatt sem. Ehelyett, és több evolúciós elméletet egyszerűsítő feltételezés ellenére18, csupán néhány száz hasznos mutáció kerülhetett kiválasztásra a csimpánzokkal lévő közös ősünk óta.19 Ez Haldane Dilemmájaként vált ismertté, amit több ellenkező véleménnyel szemben sosem oldottak meg.20 Ami ehelyett történt, az az evolúciós képzelőerő „remeke”. A hatvanas évek vége felé Kimura a semleges evolúció elméletét dolgozta ki.21 Azt gondolta, hogy a DNS legtöbb része a sejtben nem funkcionál, így szabadon tud az idők során mutálódni. Az élőlény számára tehát nem jelentkezik a működés nélküli részekhez fenntartási „költség” (a költség ebben az értelemben azt jelenti, hogy mennyivel több utódnak kell születni egy populációban, melyeket majd a természetes szelekció kigyomlál, hogy a káros mutációk eltűnjenek, illetve fennmaradjon a fitnesz22, feltéve, hogy a természetes szelekció csakis a káros mutációkat látja).
A késői ötvenes években a híres populációgenetikus, J. B. S. Haldane kimutatta, hogy a természetes szelekció nem tudja a jótékony mutációkat kiválogatni még az ember evolúciós történelme alatt sem. Ehelyett, és több evolúciós elméletet egyszerűsítő feltételezés ellenére18, csupán néhány száz hasznos mutáció kerülhetett kiválasztásra a csimpánzokkal lévő közös ősünk óta.19 Ez Haldane Dilemmájaként vált ismertté, amit több ellenkező véleménnyel szemben sosem oldottak meg.20 Ami ehelyett történt, az az evolúciós képzelőerő „remeke”. A hatvanas évek vége felé Kimura a semleges evolúció elméletét dolgozta ki.21 Azt gondolta, hogy a DNS legtöbb része a sejtben nem funkcionál, így szabadon tud az idők során mutálódni. Az élőlény számára tehát nem jelentkezik a működés nélküli részekhez fenntartási „költség” (a költség ebben az értelemben azt jelenti, hogy mennyivel több utódnak kell születni egy populációban, melyeket majd a természetes szelekció kigyomlál, hogy a káros mutációk eltűnjenek, illetve fennmaradjon a fitnesz22, feltéve, hogy a természetes szelekció csakis a káros mutációkat látja).
Négy évvel később megszületik a hulladék DNS fogalma, mely Ohno nevéhez fűződik.23 A hulladék DNS gondolata nagyon fontos az evolúciós matematika számára. Mi történne, ha kiderülne, hogy nem is létezik ilyesmi? Mi történne, ha 97% szemét helyett, a genom 97%-a működőképes?
A modern technológia mára végzett a hulladék DNS fogalmával. Ez a Humán Genom Projekt befejezése után történt. Ezelőtt is világos volt, hogy tarthatatlan volt a hulladék DNS elmélet, de sokan csökönyösen ragaszkodtak hozzá. Több okra is visszavezethető, hogy sok ember miért gondolja ma már, hogy a sejtben lévő DNS nagy része funkcionális. Például sok retrotranszpozon funkcióját felfedezték már24, amelyekről sokáig azt gondolták, hogy vírusok elemei, melyeket beinjektáltak a genomba évmilliókon keresztül.
Emellett a gének között megbúvó, fehérjét nem kódoló hosszú DNS szakaszok többségének is találtak funkciót. Úgy tűnik, hogy a genom legnagyobb része aktív. Az ENCODE projekt egy több egyetemet foglalkoztató, több millió dollárt felemésztő, több évig tartó kutatás volt, ami meghatározta, hogy az emberi genomnak mekkora része íródik át (vagyis alakul RNS-sé, ami a működés mutatója). A genomnak pusztán 1%-át analizálták, de belevonták mind a fehérjéket kódoló részeket, mind a „hulladék” területeket. A projekt kimutatta, hogy a genom bármelyik bázisa átlagosan hatféle RNS-transzkriptumot eredményezhet.25 Ez azonban messze nem azt jelenti, hogy minden fehérjévé íródik át. És azt sem jelenti, hogy mindennek kötelező funkciója van vagy hogy a betűket gyakran használják. Azt viszont igen, hogy szinte minden betű csinál valamit. Mivel általános elv a biológiában, hogy első az alak és azt követi a funkció, a tény, hogy ezek a régiók aktívak, azt sugallja, hogy van funkciójuk. Mi más miatt engedné meg a sejt, hogy olyan sok génátíródás történjen? A sejt a saját erőforrásainak tekintélyes részét arra költi, hogy fehérjét nem kódoló RNS-t hozzon létre. A sejt tehát nagyon sokat spórolna, ha kikapcsolná az ilyen energiapazarlást. A természetes szelekció az évmilliók alatt bizonyára eltüntette volna ezt az RNS-túltermelést. Azért nem tette eddig, mivel szükségesek a sejt működése szempontjából. Sőt, a genomot ma RNS-alapú számítógépnek tekinthetjük (lásd később).
A hulladék DNS hiánya egy újabb Achilles-ín az evolúciós genetika számára, mivel nélküle az evolúciós matematika nem működik, miközben egyre több funkciót fedeznek fel a fehérjét nem kódoló DNS szakaszok esetében is. Sőt, ez a „hulladék” aktívabbnak tűnik a „gének”-nél, és ezzel azt az elképzelést, hogy mi mind fehérjealapú élőlények vagyunk, a feje tetejére állítja. Egy ismert evolúció biológus, J. S. Mattick szavaival élve:
Ha nem vagyunk hajlandóak felismerni mindennek a következményeit – különösen azt a lehetőséget, hogy a nem fehérjét kódoló szekvenciák a létrejövő RNS molekulák révén párhuzamos információt közvetíthetnek –, a molekuláris biológia történetének egyik legnagyobb hibáját követhetjük el.26
A rendkívül összetett génátírás
Régebben a biológia szakos hallgatók az egy gén egy enzim elméletet tanulták. A huszadik század figyelemreméltó felfedezései alapján aztán sokan logikusan arra következtettek, hogy egy „gén” egy olyan DNS szakasz, amely egy bizonyos fehérjét kódol. A „gén” bizonyos pontnál kezdődött, és bizonyos pontnál véget ért, egyes szakaszai fehérjét kódoltak (exonok), míg talán néhány közbeeső szekvencia is volt (intronok), amelyeket kivágtak az elsődleges RNS átiratból mielőtt a „gén” fehérjévé íródott át, és emellett a gén előtti és utáni szabályozó régiókból állt, ahol különböző faktorok hozzákötődhettek a DNS-hez, a „gén” kifejeződését befolyásolva. Könnyű volt a génnek ezeket a különböző részeit felismerni. A DNS szekvenciát akár le is lehetett fordítani fehérje szekvenciává a messenger RNS szálról, ha ismerték az aminosavak három betűs kódját (ami nem bonyolult). Az „egy gén, egy enzim” elmélettel egy súlyos probléma van, hogy Darwin pángenezis elméletéhez és a hulladék DNS elmélethez hasonlóan ez is hamis.
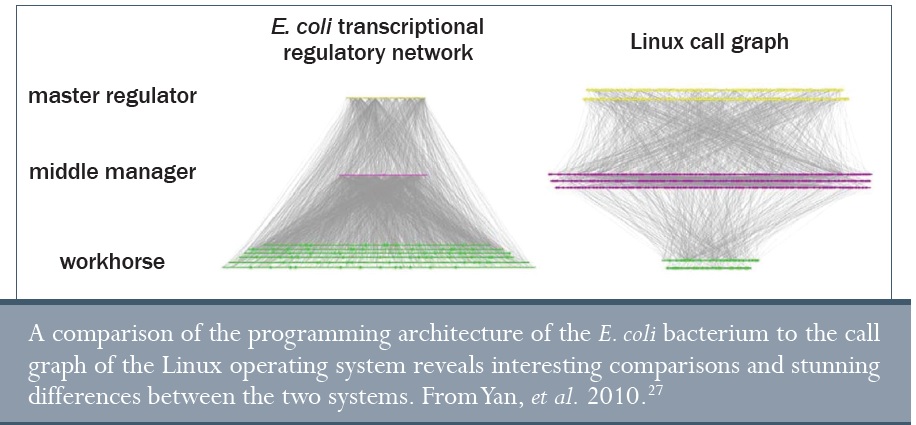
Különösen az ENCODE projekt eredményei adták meg a lehetőséget a világ számára, hogy betekintést nyerjen az univerzum legbonyolultabb számítógépes rendszerébe – az emberi genomba. De ez nem holmi fehérje számítógép. A genom inkább RNS szuperszámítógéphez hasonlít, ami fehérjét termel. A számítógéphez hasonlóan, aminek van meghajtója, újraírható memóriája (RAM), illetve képernyője, a genomikus számítógép tárolóegysége a DNS, az információs összehasonlító és számolóegysége az RNS, a végtermék pedig a fehérje.
Egyesek a genomikus szabályozó folyamatokat össze akarják hasonlítani az emberek által tervezett számítógéprendszerekkel.27 A hasonlóságok érdekesek, azonban a különbségek még látványosabbak. Például ha a Linux operációs rendszerhez hasonlítjuk, az E. coli bakteriális genomjának kevesebb magas szintű szabályozója van, amely ennek következtében kevesebb középszintű szabályozót irányít, amelyek azonban ennek következtében jóval több alacsony szintű kimenetet határoznak meg. Úgy tűnik, mint ha a genom úgy volna optimalizálva, hogy a leghatékonyabb módon működjön. A Linux helyett azonban lehet, hogy jobb összehasonlítani a hadsereg számítógépeivel, amelyeknek nagyon rövid programjaik vannak, jellegzetesen minimális parancskészlettel. Ez azért van, mert korlátolt számú parancsot hajtanak végre a leghatékonyabb módon ahelyett, hogy több dolgot egyszerre hajtanának végre (mint például grafika, játékok, zene, illetve szövegszerkesztés egyszerre, amit a Linux megenged). De van itt még egy rejtély: rengeteg munkaórába telt azokat a számítógépeket megtervezni, amelyek a B-52-es bombázókat vezérlik, de egyetlen hiba ezekben a programokban vagy az alrendszerekben katasztrofális összeomlást okozhat abban a rendszerben, amelyet szabályoznak. Az emberi genom ehhez képest azonban sokkal komplexebb, és több ezer hibát is el tud bírni anélkül, hogy összeomlana.
Több dolgot végez egyszerre, és jobban van megtervezve!
Amikor az emberi genom szekvenálását befejezték, a kutatók meglepődtek, amikor mindössze 23,000 „gént” találtak benne.28 Már tudtuk, hogy sokkal több ennél az emberi testben lévő fehérjék száma. Ez hogy lehetséges? Néhány évvel később az ENCODE projekt olyan nyomokra bukkant, amelyek arra utaltak, hogy az emberi genomban nagymértékű alternatív splicing megy végbe.29 Ahogyan már láttuk, a „gén” minden egyes részét többféle fehérjéhez fel lehet használni.30 A test valahogyan tudja, hogyan kell létrehozni különféle fehérjekombinációkat külön fehérjekódoló egységekből, és úgy összeállítani őket a splicing folyamata során, hogy azzal több százezer egyedi fehérje jöjjön létre. Sőt mi több, más-más sejttípus különböző fehérjéket is elő tud állítani ebből a folyamatból. Emellett különböző fehérjék más-más időpontokban állítódnak elő, így a sejtek azt is tudják, hogy mit, mikor, és milyen körülmények között kell előállítani.31 Valami irányítja ezt a folyamatot, és nem biztos, hogy a genom fehérjetartalmú része.32
Minden egyes „génbe” be van ágyazva több kisebb kód. Mindegyik pusztán néhány betű hosszúságú, de több kód előfordul minden exon és intron elején és hátulján. Ezekből áll össze az ún. spliceoszóma, ami a genomnak az a része, ami az exonok kombinálódását, vagyis a gén splicing-nak nevezett folyamatot szabályozza. A spliceoszóma összetettsége, illetve az eukarióták egész genomjának komplexitása még egy Achilles-ín az evolúciós genetika számára. A genom túl bonyolult, és ez a mutációs „céltábla” túlságosan nagy ahhoz, hogy az ismert fajok évmilliókig túléljenek33, nemhogy előtte kialakuljanak.
Lehet, hogy az olvasó figyelmét felkeltettem azzal, hogy a „gén” szót idézőjellel használom ebben a fejezetben. Ez azért van, mert már nincsen egyértelmű definíciója a szónak.34 Legalább is, a definíció annyira megváltozott, hogy valami teljesen mást jelent minden, a baktériumoknál bonyolultabb élőlényeknél. A genetika egyre bonyolultabb lesz, és az egyszerű, régi elképzelés már idejét múlt. Ezért innentől kezdve, amikor a „gén” szó előfordul, vegyük a klasszikus értelemben: a gén egy olyan DNS szakasz, ami fehérjét kódol. Ennek a definíciónak csak az a problémája, hogy egy bizonyos DNS szakasz többféle fehérjében használódhat fel, kontextustól függően.
A négydimenziós genom hiper-komplexitása
Fokozzuk még egy kicsit a dolgot, és tekintsük át a komplexitás egy másik szintjét. Amikor az emberi genom szekvenálása sikeresen megtörtént, azt hittük, hogy ettől érteni is fogjuk, hogyan működik. Ez naiv elképzelés volt. Amit elértek, az pusztán a nukleotidok egy lineáris szekvenciájának meghatározása volt. Ez egy legalább négy dimenzióban működő genom első dimenziója. Ez mit jelent? Gondoljunk a DNS-re. Egy szakasz, egy vonalszerű valami, ami definíció szerint egydimenziós. Amikor az emberi genomot befejezték, a hulladék DNS elméletet azt támasztotta alá, hogy a gének szét vannak szórva a genomban, és a rokon funkciók nem csoportosulnak össze. „Nahát!”, mondhatták, „a gén sorrend random jellegű, ami a véletlenszerű, időbeli változások eredménye.” Ez rövidlátó megállapítás volt, hiszen csak az első dimenziót figyelték meg.
Épp az imént hallottunk a splicing folyamatról. Itt a genom egyik része hatással van a másik részre, vagy közvetlenül, vagy az RNS és/vagy fehérje közvetítésével. Ez a rész a genom második dimenziója. Ahhoz, hogy ezeket a kölcsönhatásokat felvázolják, a genomot le kellene rajzolni, és nagyon sok nyilat kellene húzni egyik pontból a másikba. Ehhez több papírlapot kellene használni, amelynek két dimenziója van (magasság és szélesség).

A genom második dimenziója rendkívül bonyolult, és számos eleme van – egyebek között specifitási és transzkripciós faktorok, enhancerek (erősítők), aktivátorok (regulátor gén), represszorok (a regulátor gén fehérjeterméke), hiszton (kromoszómafehérje típus) acetiláz valamint DNS metilációs jelek, az RNS poszt-transzkripciós szabályozása, alternatív splicing. Fő szerepe azon események koordinálása és szabályozása, amelyek mind a sejtmagban, illetve a sejten belül is lejátszódnak. Ebben a dimenzióban a sorrend nem nagyon fontos, mivel a génszabályozóknak így is összevissza lebegniük kell, hogy megtalálják a célpontjukat. Nem fontos, hogy a célpont épp a szabályozó mellett legyen. A következő szint azonban nagyon érdekes lesz.
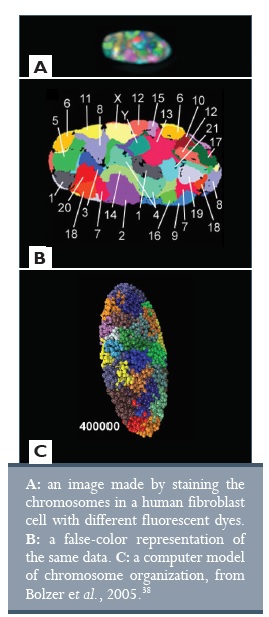
A genom harmadik dimenziója a DNS háromdimenziós alakja a sejtmagban. Ezen a szinten a gének nincsenek véletlenszerűen szétszórva a genomban, hanem szükség szerint sorba rendezve. A sorba rendezett gének nem feltétlenül találhatók meg egymás mellett a kromoszómákon, viszont amikor a kromoszómák összehajlanak, ezek a gének egymás mellé kerülnek a háromdimenziós térben, és gyakran egy sejtmagmembránba ágyazott pórus mellett vagy a transzkripció közelében csoportosulnak össze.35
Valami azonban mégis rögzíti a helyüket. Mivel a DNS egy óriási összegyömöszölt cérnára hasonlít, ahol a belső részeket nehéz elérni, míg a kívülre eső részeket könnyebb elérni.36,37
Az első dimenzióba ágyazott kód egy része meghatározza a DNS háromdimenziós „hajtogatását”, így a génexpressziós mintázatokra is hatással van. Ez a harmadik dimenzió kiemelkedően fontos.38
 A genom negyedik dimenziója az első, második és harmadik dimenziókban történő időbeli változásokat foglalja magába. A kromoszómák különleges alakot vesznek fel a sejtmagban, de ez az alak az egyedfejlődés során megváltozik, mivel a különféle sejttípusoknak többféle génösszetételre és más genetikai végrehajtási parancsokra van szükségük. Az alak rövidtávon is meg tud változni, ahogy a sejtek különféle ingerekre válaszolnak, illetve a DNS különböző részeit is kitekerik, hogy a mélyben fekvő génekhez jussanak, majd visszacsomagolják, ha már nincsen szükség az adott génre.39
A genom negyedik dimenziója az első, második és harmadik dimenziókban történő időbeli változásokat foglalja magába. A kromoszómák különleges alakot vesznek fel a sejtmagban, de ez az alak az egyedfejlődés során megváltozik, mivel a különféle sejttípusoknak többféle génösszetételre és más genetikai végrehajtási parancsokra van szükségük. Az alak rövidtávon is meg tud változni, ahogy a sejtek különféle ingerekre válaszolnak, illetve a DNS különböző részeit is kitekerik, hogy a mélyben fekvő génekhez jussanak, majd visszacsomagolják, ha már nincsen szükség az adott génre.39
Még a „hulladék” DNS-ben is történnek változások. Például nagymértékű retrotranszpozon (ugráló gén) aktivitás történik az agy fejlődése során, amikor több osztálybeli DNS (L1, Alu illetve SVA) lemásolják magukat, és az egyedi agysejtekben lévő genomokon belül összevissza mozognak. Ez a különféle agysejtek differenciálódását segíti elő.40 Hasonlóan a májsejtek is számos kromoszóma duplikációval rendelkeznek. A test különböző sejtjeinek végső genomja nem feltétlenül ugyanaz, mint ami a megtermékenyített petesejtben volt, mielőtt az elkezdett volna osztódni, ugyanakkor a genom alakja sejtek között és időben is változik. Ezek a példák nem a véletlen művei, hanem a genom változásainak gondosan ellenőrzött, négy dimenziós szimfóniája. Remélem, sikerült lenyűgöznünk, hisz
a genomot egy mesteri Építész készítette.
Ez mind nagyon bonyolult, de fontos előzmény az evolúciós genetika következő Achilles-inának megértéséhez. A darwinizmusnak az a jó, ha az élet egyszerű. A természetes szelekciónak szüksége van arra a képességre, hogy felismerje a mutációk okozta kis változásokat, és egy állatcsordából a legéletképesebb egyedeket kiválassza. Amikor egy faj már létezik, a természetes szelekció korlátozott módon tud dolgozni (lásd előző fejezetet), de ez a folyamat meg tudná magyarázni azt, hogy hogyan is keletkeztek a fajok? Aligha, hiszen egy hibafelhalmozó és kiválasztó rendszer nem tud létrehozni komplex, egymással kölcsönhatásban lévő, négydimenziós rendszereket, amelyek hatalmas mennyiségű adatot képesek tömöríteni, emellett végtelenül rugalmasak. És ha már létezik is ez a rendszer, komoly fenyegetést jelentenek számára a random mutációk okozta változások. Ma ez a helyzet. Rendben van, hogy apró változásokat képzelünk el egy már létező, bonyolult rendszerben. Viszont ha ezeket a kis változásokat arra használjuk fel, hogy magának a rendszernek az eredetét megmagyarázzuk, akkor ez olyan, mintha azt mondanánk, hogy a legújabb számítógépes operációs rendszer kifejlesztésében semmilyen intelligencia nem vett részt. Pedig a genom a ma létező összes operációs rendszert felülmúlja mind komplexitásban, mind hatékonyságban.
A kodon degenerálódása
Ma az emberek által tervezett számítógépek a két bites rendszer szerint működnek, mivel a tranzisztorok csak két betűt kezelnek, amivel dolgozni tudunk (0 és 1, nincs és van). Az ebből származó matematika elég egyszerű, mégis bonyolult számítógépes chipeket állítottunk elő, amelyek ezen az elven működnek. A genom azonban négy bites rendszeren alapszik. A 0 és az 1 helyett négy betű van a genomban (A, C, G és T). Ez nem lenne szükségszerű, hiszen ugyanazt az információt ki lehet betűzni bármilyen számú betűvel. Miért pont négy bit?
Furcsa módon, három betűre van szükség egyetlen aminosav kódolásához. A genomban ezek a hárombetűs kodonok csoportokba rendeződnek (ún. exonok), ahol minden csoport egy vagy több fehérje részleges funkcióját kódolja. Összesen 20 aminosav van az emberi fehérjékben, de 64 lehetséges kodon létezik (három pozíció és négyféle betű szerint 4 x 4 x 4 = 64 kodon). Ez azt jelenti, hogy néhány aminosav, mint például az alanin, több kodonhoz rendelhető (GCA, GCC, CGC és GCT). Más aminosavak, mint a triptofán csak egy kodonnal rendelkeznek (TGG).
Látható, hogy az alaninnak mind a négy kodonja „GC” betűkkel kezdődik. Ez azt jelenti, hogy bármely mutáció, ami az utolsó betűt változtatja meg, ugyanazt az aminosavat eredményezi, noha a kodon más. Ezt kodon degenerálódásnak hívjuk és a genom ellenállóságát növeli, mivel így néhány mutáció a fehérjekódoló régiókban alig okoz változást. Alig, mert a többféle kodonhoz tartozó transzfer RNS nem ugyanazzal a gyakorisággal fordul elő. Vannak példák, amikor egyetlen betű megváltozik, és noha az aminosav nem változott meg, mégis egy káros mutáció jön létre. A fehérje átíródása közben volt egy szünet, amíg a ritka tRNS előkerült. Emiatt aztán a fehérje rosszul tekeredett, az eredmény pedig egy hibás enzim. Több új megfigyelés szerint a fehérje átíródás általában a kodon használatától függ, legalábbis a baktériumokban, ahol az átíródás sebessége a megfelelő kodonok alkalmazásán múlik. A bakteriális sejtek génjeiben általában nincs promoter. Amikor mégis előfordulnak, az átíródás sokszor ideiglenesen lelassul, mivel a bakteriális riboszóma ragaszkodik a promoter szekvenciákhoz.41 Így az alternatív kodonokkal, a Tervező mérnöki pontossággal hozta létre a géneket, hogy kevesebb belső ellentmondás jöjjön létre más genetikai tulajdonságokkal.
Mindezek mellett nagyon jó oka van a kodon degenerálódásnak, illetve a négy bázis alapú genomnak. Ez nem csupán a 20 aminosav kódolásának legoptimálisabb módja42, de több egymással átfedésben lévő kódot is lehetővé tesz.
A többszörösen átfedő kódok
Minden magasabb rendű szervezet genomja meglehetősen nagy mennyiségű adatot tömörít. Az ilyen genomok adott szakasza több dolgot tud párhuzamosan végezni. A DNS-nek egyetlen betűje egy exon része lehet, amely azonban húszféle fehérjéhez is felhasználódhat. Ugyanakkor az a betű része lehet a splicing kódnak, amely megmondja a sejtnek, hogy mikor hozza létre azokat a fehérjéket. Az a betű a hiszton kód része is lehet, amit a sejt arra használ, hogy megtalálja azt a helyet, ahol a hisztonokat (védő fehérjéket) a DNS köré tekerje. Az a betű még a DNS háromdimenziós alakját is befolyásolhatja. A kiterjedt epigenetikai kód része is lehet (lásd első fejezetet), és lehet, hogy része egy hárombetűs kodonnak amelyet egy bizonyos aminosavra fordít a rendszer. A genom négy bázisú alapjának és a kodon degenerálódásnak köszönhetően a Tervező képes volt többféle kód közül választani, amikor több egyidejű követelménynek kellett megfelelni. A degenerálódás megengedi azt is, hogy több genomikai parancs átfedésben legyen anélkül, hogy a fehérjék létrehozásának feltételeit veszélyeztetné.
A többszörös, átfedő DNS és RNS kódok ellentmondanak a naturalista magyarázatoknak, és lehetetlenné teszik a természetes szelekció számára, hogy a hosszú távú evolúciós változásokat létrehozza. A szelekció a „nehézségek felülkerekedhetetlen falába” ütközik, amikor olyan mutációkkal áll szemben, amelyek egyszerre több tulajdonságot is érintenek. A polifunkcionalitás (más néven pleiotrópia) azt jelenti, hogy egy adott mutáció egymástól teljesen független tulajdonságokat is érinthet (pl. színlátás, a fokhagyma szagának elviselése, a mitokondriális hatékonyság, noha ez utóbbi extrém példa lenne). Ez az evolúciós genetika újabb Achilles-ina. Hogy tud egy próbálkozásokon alapuló rendszer, amely mindig a legegyszerűbb választ keresi egy környezeti problémára, egy többszörösen átfedő szabályozási rendszert létrehozni? Egy ilyen rendszer a világegyetem nagy csodája lenne. A „multitasking”nélkül a genomnak sokkal nagyobbnak kellene lennie, másrészt a DNS-alapú többsejtes élőlények sem tudnának nélküle létezni.
A teremtés bizonyítéka a génjeinkben
A modern genetika egy hihetetlenül bonyolult világot tárt fel, de ez nem minden, mivel a genetika révén történelmi elméleteket is meg tudunk vizsgálni. Meglepné, hogy a sok különböző teremtés elbeszélés közül az egyik különösen egybecseng azzal, amit a genetikából tudunk? Meglepné, ha megtudná a teremtésről, az özönvízről, illetve a bábeli torony történetéről, hogy bőséges bizonyítékokat találunk rájuk a génjeinkben?
Mivel Mózes első könyve, a Genezis, egy történeti könyv, és mivel az emberiség történelmének kezdeteiről tesz tanúbizonyságot, bizonyos konkrét előrejelzéseket is ad az emberi genetikával kapcsolatban. Ezek az előrejelzések abból indulnak ki, hogy eredetileg két embert (Ádámot és Évát) teremtett Isten, illetve hogy mintegy 1,600 évvel később (Noé idejében) egy populáció beszűkülés történt, amikor az emberiség létszáma nyolcra csökkent, majd néhány száz év múltán a Közép-Kelet egy központi helyéről elindult az emberiség szétterjedése. Óvatosan kell ehhez a témához közelíteni, mivel az ember által művelt tudomány sokszor elég rosszul derítette ki az igazságot az évszázadok alatt. Ma is elég sok mindenben tévedhetünk.
Ennek ellenére, ha figyelmesen megvizsgáljuk a rendelkezésre álló adatokat, könnyen megláthatók a bibliai történet bizonyítékai a génjeinkben. Amikor olyan emberek, mint Francis Collins, aki a Humán Genom Projekt vezetője, és állítólagos evangéliumi keresztyén, azt állítják, hogy Ádám és Éva létezésére nincsen bizonyíték,43 akkor arra is kevés a bizonyíték, hogy ezek az emberek vajon elgondolkodtak-e azon, hogy a Biblia mit is mond az emberi eredetről.
A Biblia azt állítja, hogy az egész emberiség egyetlen pártól származik. Ez egy mély értelmű állítás a genetikáról, mivel nagyon bekorlátozza azt a humángenetikai diverzitást, amit ma látunk. De ne ragadjunk le az evolúciós gondolkodásmódnál, ami csak a végtelen időbe visszanyúló modelleket tudja elfogadni. Minden Ádám és Éva után következő embernek kapcsolódnia kellett hozzájuk szexuális rekombináció és szaporodás révén, de ez nem szükségszerűen igaz magára az alapító emberpárra. Isten többféle ivarsejt-vonalat beleteremthetett Ádám hereszövetébe. Hasonlóan, Éva petesejtjeit is több, egymástól teljesen eltérő emberi genommal tervezhette meg. A legkorábbi emberi populációban nagyon nagy diverzitás lehetett. Másként lehet, hogy csak egyetlen genomból származunk, és Éva pedig egy közeli klónja (az Y kromoszóma nélkül) vagy akár haploid klónja is lehetett Ádámnak. Így az emberi genetikáról szóló bibliai modellek sok lehetőséget állítanak elénk. Melyiket válasszuk?
Teológiai okok miatt arra gondolnék, hogy Éva Ádám klónja. Így Éva is az Ádámra és utódaira kimondott isteni átok alá került, de egyben Krisztus családjának tagja, aki vérrokon Megváltónk.44 Emellett, mivel Ádám oldalbordájából formálódott, akkor azt is feltételezhetjük, hogy Isten az Ádámból származó sejteket, izmokat, idegeket, véredényeket, illetve DNS-t (!) felhasználta, mikor Évát megalkotta. A Biblia ezt nem mondja ki egyedül érvényesként, és más lehetőségek is vannak, de én inkább erre hajlok.
Lehet, hogy nagyon meglepődsz ezen, különösen, ha Francis Collins-t meghallgattad, vagy olvastad az írásait, de a ma megfigyelhető rengeteg genetikai diverzitás teljes mértékben beleférhetett Ádámba és Évába. Sőt, a legnagyobb része egyedül Ádámba is beleférhetett! Az emberi genomban összesen 10,000,000 helyen fordulnak elő az általános jellemzők. Az átlagembernek ezekből két-három millió van. Más szavakkal, körülbelül három millió olyan pozíció van, ahol a testvér kromoszómák (a kromoszómák párosával vannak; így például két példányod van az 1. kromoszómából) más betűt tartalmaznak ugyanazon a helyen. Nem nagy dolog elképzelni, hogy Ádám ezeket a változatokat majdnem mind a genomjában hordozta. Az emberek miért hordoznak az összesből csak egy töredéket? Ez valószínűleg a családfájukban történő rekurzió (önhasonló sokszorozás) miatt van. Vagyis, a genomból azonos részeket örököltek ugyanabból a távoli őstől különböző vonalakon keresztül. Ehhez még vegyük hozzá a populáció növekedést, összezsugorodást, feloszlást, és a keveredést a bibliai időkön keresztül45, és látni fogjuk, hogy nincs szükség több millió emberre, akik az ősidőkben éltek, hogy megmagyarázzuk a jelenlegi humángenetikai diverzitást.
Mi több, a legtöbb közönséges genomvariáns két példányban van jelen. Amikor ennél több változatot találunk, ezt egy korai mutációval lehet megmagyarázni (pl. a 0-s vércsoport leggyakoribb változata egyértelműen az A vércsoport allélját kódoló gén mutációja [hibás másolata]46, miközben világszerte megtalálható), vagy egy Bábel után történt mutációról lehet szó (pl. a sarlósejtes gén, amely Afrika egyes részein megtalálható, vagy a kék szemért felelős gén, amely Észak Európában honos, vagy bizonyos immunrendszeri gének sok változata, amelyek arra vannak tervezve, hogy gyorsan megváltozzanak). Így, mivel a variációk nagy része beleférhet egyetlen emberbe, talán nincs is szükség a furcsa kora-bibliai genetikai modellekre.
Az afrikai származásról szóló elmélet esetén („Out of Africa” modell) az evolucionisták szerint az emberiség majdnem kihalt, amikor a genetikai diverzitásunk nagyon beszűkült, majd ezután kiterjedt, és előbb-utóbb elhagyta Afrikát. A beszűkülés miért része a modellnek? Mivel azt akarták elmagyarázni, hogy az emberi diverzitás hiánya hogy terjedt szét a világban.47 Ez a diverzitás sokkal kisebb, mint amit a kezdeti feltételezések alapján az Afrikában mintegy egymillió évig élő nagy létszámú populációk kiadtak volna. A beszűkülés az evolúciós elmélethez csapott utólagos kiegészítés, de az alacsony diverzitás mindig is része volt a teremtés modellnek.
Mellesleg, érdekes módon, az állatok másmilyenek. Az emberek meglepően egyöntetűek az állatfajokhoz képest. A csimpánzok ötször vagy hatszor annyira változatosak. A közönséges házi egér populációi nagyfokú variációt mutatnak, beleértve többszörös kromoszóma átrendeződéseket. Ilyen példák gyakoriak az állatvilágban. Ez olykor az állatok genomjának időbeli degradációja miatt van. Attól is lehet, hogy az állatok más-más pontból indultak az emberhez képest. A Biblia nem azt mondja, hogy minden állatból kettőt-kettőt teremtett. Sőt, azt is feltételezhetjük, hogy egy világméretű, funkcionáló ökoszisztémát hozott létre, ahol minden fajta élőlényben magas fokú diverzitás létezett (amit a fosszilis leletanyagban is lehet látni). Az azonban igaz, hogy a legtöbb állatból csak két-két példány ment be Noé bárkájába, de ezek egy potenciálisan sokkal nagyobb genetikai állományból kerültek elő. Mivel a legtöbb genomikai adat ma az emberből származik (világos okok miatt), messze vagyunk még attól, hogy a legtöbb állatfajra genetikai modellt alkossunk, de több érdekes dolgot is felvethetünk.
Az emberi genomban előforduló általános változatok mellett számtalan ritka variáns is létezik. Ezek csak elszigetelt embercsoportoknál fordulnak elő, és van arra utalás, hogy ezek olyan mutációk, amelyek Ádám, vagyis inkább Bábel óta léptek fel. Ebbe beletartozik az európaiak kék szeme, a sarlósejtes vérszegénység Afrikában, illetve a nagyon ritka variánsok, amelyek csakis egyetlen törzsben, családban, vagy emberben fordulnak elő. Nagyon sok genetikai változaton osztozunk. Ez arra utal, hogy a közelmúltban létezett kisméretű populációtól származunk. Ami azonban nem egységes bennünk (populáció szintű variáció gyanánt) az arra mutat, hogy a genomunk világszerte gyorsan hanyatlik, elég gyors mutációs rátával, minden embercsoportban. Erről később még lesz szó.
Amikor a nőkre, illetve férfiakra specifikus DNS részeket megtekintjük, egyre több bizonyítékot látunk Ádámra és Évára. Sok kísérleti bizonyítékkal rendelkező elmélet szerint a mitokondriumok (a sejten belüli kisméretű energiatermelő sejtszervecskék, amelyek a cukrot energiává alakítják át) csak a női vonalon adódnak tovább. Mivel a mitokondriumnak is megvan az 16,569 betűből álló kis genomja, és mivel ez a kis genom szintén mutációkon megy keresztül az idő során, egy világméretű női családfát tudunk a segítségével felvázolni. Ez az afrikai mitokondriális Éva elméletéhez vezetett az evolúciós irodalomban 1987-ben.48 Ha több más nő élt volna évmilliókkal ezelőtt, csakis egynek sikerült átadni a genomját a ma élő embereknek.
A mitokondriális Éva létezését több százezer évvel ezelőttre teszik, de csak akkor, ha nagyon lassú mutációs rátát, valamint a csimpánzokkal lévő közös eredetet feltételeznek. Ha azonban a valós mutációs rátát alkalmazzuk, Éva kora 6,000 évre jön ki.49 Egy nemrég megjelent tanulmány szerint a mitokondrium kontroll régiójában lévő mutációk (amelyek a mitokondriális mutációk kétharmadát teszik ki) minden második generációban lépnek fel.50 Mivel Éva mitokondriális szekvenciáját meghatározták és leközölték az evolúciós szakirodalomban51, és mivel a mitokondriális vonalak kevesebb, mint 30 mutációra vannak az „Éva” konszenzus szekvenciától, és mivel a legtávolabbiak csak 100 mutációra vannak tőle, az emberi populációk mitokondriális DNS-diverzitása belefér a 6,000 éves/200 generációs időskálába.
Az afrikai Éva elméletről és az evolúciós feltételezésekről már korábban szóltunk52, így a részleteket nem ismételjük meg itt. Az eredeti nő mitokondriális genomjának több mint 99%-át ismerjük. Másról nincs bizonyíték. Miért hiszik el oly sokan, hogy Éva csak egy nő volt egy nagy populációban, amely nagyon régen élt? Azért, mert ez az evolúciós történet része. Nem a valóságon alapszik, de könnyű kifogást kínál nekik, hogy ezzel a bibliai előrejelzéseket figyelmen kívül hagyják, miszerint csak egy női vonal létezik világszerte.
A mitokondriális genomhoz hasonlóan a férfiak Y kromoszómája is lehetőséget nyújt, hogy egy férfiakra alapuló családfát szerkesszünk. Feltételezések szerint az Y kromoszómájú Ádám is nagyon sok ideje élt, de más időben, mint a mitokondriális Éva. Ám, ahogy fent láttuk, ezek a végkövetkeztetések olyan modelleken alapulnak, amelyek nagyon sok dolgot feltételeznek az emberi történelemről, a populáció méretekről, illetve a mutációs rátáról. A csimpánz Y kromoszómájának a javított verziójával53, és azzal a felfedezéssel, hogy csak 70%-ban azonos az emberi Y kromoszómával (még ez a szám is nagyvonalú, mivel a csimpánz Y kromoszómájának a fele hiányzik), az evolucionisták arra kényszerülnek, hogy azt mondják, hogy az Y kromoszóma rettenetesen gyorsan mutálódott az emberi történelem során. Azonban a jelenlegi emberi Y kromoszóma szekvenciák mind nagyon hasonlítanak egymásra. Az egyetlen módja annak, hogy magas fokú hasonlóság létezzen magas mutációs ráta mellett az az, ha nemrég élt a közös ősünk. Éljen Ádám!
Egy másik módszer, hogy Ádám és Éva valódiságát bizonyítsuk, a kapcsoltsági adatok. A szexuális szaporodás során mindkét szülőben lévő sejtek átmennek a meiózison, ahol a szülőktől örökölt kromoszómák összekeverednek. Így amikor a szülők átadják a génjeiket, valójában a nagyszülőktől örökölt kromoszómák összekevert változatait adják tovább. Ez a fajta összekeveredés, amit crossing overnek (reciprok kicserélődés) neveznek, azt eredményezi, hogy a DNS nagy tömbökben örökítődik át. A genomnak vannak részei, amelyek az emberi történelem során sosem cserélődtek ki (ami egy fiatal genomot feltételez). Amikor a két variánst együtt öröklik (mivel közel vannak egymáshoz ugyanazon a DNS szálon), akkor kapcsoltnak hívják. A kapcsoltságot részleteiben vizsgálták, és elég sok érdekes dolgot tanultunk róla. Először is, kettő-négy tömb határozza meg az emberi populációkban lévő tömbök többségét.54 Más szavakkal, csak néhány eredeti kromoszóma létezik, és azoknak a darabjai még érintetlenek. A genomban mindössze tízezer körüli blokk van, ami könnyen megmagyarázható, ha az emberi populáció pusztán 200 generációt élt meg, és a hosszabb kromoszómakarok csak egy vagy kétszer cserélődtek karonként és generációnként.
A legtöbb evolúciós tanulmány mögött lévő molekuláris órára alapuló feltevések mellett a legtöbb elmélet azzal számol, hogy a rekombináció ugyanúgy megy végbe minden időben és földrajzi helyen. Ez azonban nem feltétlenül van így, mivel az kicserélődést több genetikai tényező is befolyásolja (pl. a PRDM9 gén esetében különösen), és hogy ezeknek a faktoroknak variánsai vannak, amelyek a kicserélődés sebességét befolyásolják a különböző egyedekben.55 Ez kihívást jelent sok előzetes tanulmány számára, beleértve azokat az elméleteket, amelyek az ember afrikai eredetéről szólnak.
Afrikából vagy Bábelből származunk?
A fentiekben az ember afrikai eredetéről beszéltünk a mitokondriális Évával kapcsolatban. Ebben a részben összehasonlítjuk az „afrikai eredet” és a Teremtés közös elemeit. A leggyakoribb evolúciós értelmezések szerint mindannyian egy kis populációból származunk, amely sok kisebb csoportra oszlott egyetlen esemény hatására, ami után az emberiség az egész világra szétterjedt. Ez a szétterjedés a közelmúltban történt három fő női vonallal és egy fő férfi vonallal. Ja, és a Közel-Keleten haladt át, mielőtt Európába, Ázsiába, Ausztráliába, Óceániába, vagy a két amerikai kontinensre érkezett. Ezeket a pontokat a Teremtés könyvében leírt özönvíz elbeszélés (1.Mózes 6-8), a Bábel tornya (1.Mózes 11), illetve a nemzetség táblák (1.Mózes 9-11) mind elbeszélik. A különbség az időzítésben (6.000, 4.500, illetve kb. 4.000 évvel ezelőtt, szemben a több tízezer évvel ezelőtti időszakhoz képest), illetve az eredet földrajzi helyében van (Közel-Kelet, illetve Északkelet-Afrika a Vörös tengernél56).
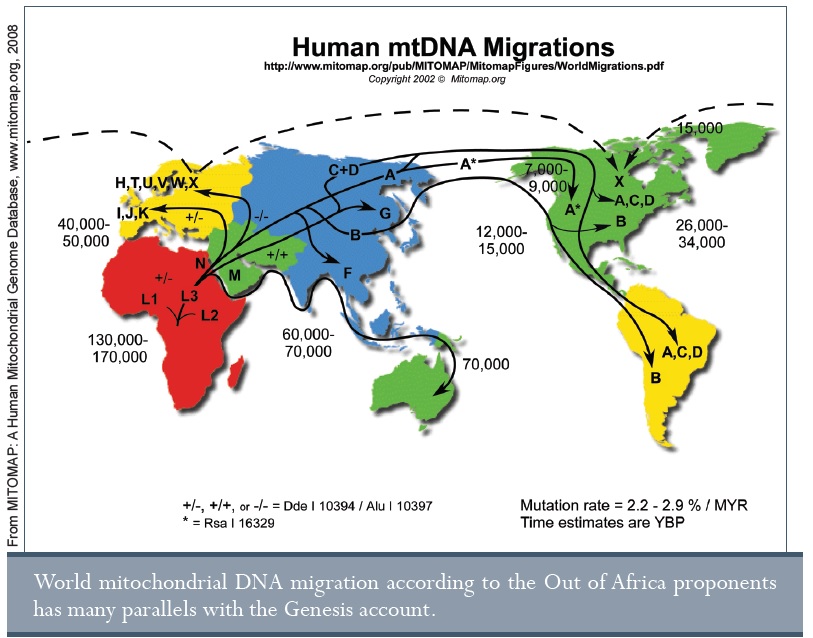
Az afrikai eredetmodell következtetéseit azonban az evolúciós alapfeltevések irányítják.57 Valójában egy molekuláris órát feltételeznek, ami az idők során a mutációkat az összes embercsoportban egyforma sebességgel halmozza fel. Már láttuk, hogy ez nem igaz. Azt feltételezik, hogy az összes emberi populáció hasonló demográfiai paraméterekkel rendelkezik (ugyanaz a születési és halálozási ráta, átlagos házassági életkor, átlagos utódszám, stb.). Azt is feltételezik, hogy nincsen különbség a populációk között a DNS javítómechanizmusokban, mivel ez a molekuláris órákat összezavarná. Így, amikor nagyobb diverzitást fedeznek fel Afrikában, automatikusan azt a következtetést vonják le, hogy ez egy idősebb populáció, vagyis az egész emberiség forrása. De mi van, ha egyes afrikai törzseknek más a történelmük, mint a többi törzsnek? Sok olyan afrikai van, aki belefér az átlagos mitokondriális szekvencia készletbe. Mások pedig nagyon különböznek. Ez azt jelenti, hogy ezek öregebb szekvenciák, vagy hogy ezek több mutációt szedtek össze a mitokondriális vonalukban valamilyen oknál fogva? Érdekes módon, egy új tanulmány azt állította, hogy a korai afrikai törzsek évezredeken keresztül kicsik maradtak és egymástól el voltak szigetelve.58 Ez gyors mutációs felhalmozódáshoz, illetve genetikai sodródáshoz vezet.
Az afrikai eredetmodell és a teremtéstörténet között még találunk párhuzamokat. Általában azt gondolják, hogy az emberek Afrikában éltek több millió éven keresztül, mint pl. a Homo erectus. Azután pedig ezek a modern ember előtti populációk váratlanul összeomlottak, és majdnem kihaltak. A tízezer vagy annál kevesebb túlélő pedig valahogyan életben maradt, majd gyorsan modern emberré fejlődött. A populációk újra felszaporodtak, sokfélék lettek, és néhány genetikai vonal Afrikából kikerülve az egész világon elterjedt.
 A Biblia szerint kezdetben két ember volt, Ádám és Éva. A populáció ismeretlen számig növekszik, majd összeomlik, lecsökken nyolc emberre, három szaporodóképes párra, az özönvíznél, 1,600 évvel a teremtés után. Az emberek létszáma ezek után újra növekszik, de továbbra is ellenállnak Istennek, mint az özönvíz előtt, így Ő beavatkozik, és összezavarja a nyelvüket Bábelnél, aminek következtében törzsenként eltérő útvonalon indulnak el. Így néhány generációval az özönvíz után szétterjednek, hogy a világot benépesítsék a bábeli zűrzavar után. Vannak párhuzamok az afrikai eredetmodell és a teremtéstörténet között, ha az evolúciós elméletet az meglévő eredményekhez adaptáljuk. Az afrikai eredetmodellnek különösen a világszerte tapasztalható alacsony genetikai változatossággal kell megbirkóznia (vagyis a populáció-összeomlással), valamint az emberiség egyszeri szétterjedésével, ami a génjeinkben látható.
A Biblia szerint kezdetben két ember volt, Ádám és Éva. A populáció ismeretlen számig növekszik, majd összeomlik, lecsökken nyolc emberre, három szaporodóképes párra, az özönvíznél, 1,600 évvel a teremtés után. Az emberek létszáma ezek után újra növekszik, de továbbra is ellenállnak Istennek, mint az özönvíz előtt, így Ő beavatkozik, és összezavarja a nyelvüket Bábelnél, aminek következtében törzsenként eltérő útvonalon indulnak el. Így néhány generációval az özönvíz után szétterjednek, hogy a világot benépesítsék a bábeli zűrzavar után. Vannak párhuzamok az afrikai eredetmodell és a teremtéstörténet között, ha az evolúciós elméletet az meglévő eredményekhez adaptáljuk. Az afrikai eredetmodellnek különösen a világszerte tapasztalható alacsony genetikai változatossággal kell megbirkóznia (vagyis a populáció-összeomlással), valamint az emberiség egyszeri szétterjedésével, ami a génjeinkben látható.
A meglepő neandervölgyi
Mit tegyünk az állítólagos nem emberi barlanglakó őseinkkel? Újabb régészeti és genetikai tanulmányok hatására a neandervölgyiekről alkotott evolúciós elgondolások az elmúlt évtizedben nagyon sokat változtak. A neandervölgyiekről most az az elképzelés, hogy barlangokban festettek, zeneszerszámokat készítettek, szabályozott módon használták a tüzet, a halottaikat rituális módon temették el fejükkel a nap irányában, és különféle ritka ásványokat gyűjtöttek össze azért, hogy összeporítsák és kozmetikumot készítsenek belőlük.59 Ezeknek a részleteit még vitatják, de akárcsak egy ilyen gondolat, főként egyszerre több említése, néhány évvel ezelőtt evolúciós eretnekségnek számított volna.
A fejlődő technológiának köszönhetően most képesek vagyunk arra, hogy a DNS-t kivonjuk a legjobban konzervált neandervölgyi csontokból is. Az ősi DNS kutatás azért problematikus, mivel a DNS törékeny molekulája könnyen szétbomlik az egyed halála után. Ráadásul a károsodás hasonló ahhoz, ami az egyed élete során történik. Így nehéz különbséget tenni az egyed halála után bekövetkezett DNS károsodás és az egyed felmenőiben végbement mutációk között. Egy másik probléma a szennyeződés. Mivel az ősi mintában lévő DNS szükségszerűen degradálódik, bármilyen mai DNS-sel történő szennyeződés elnyomja az ősi DNS bizonyítékait. A kutatók jól ismerik ezeket a problémákat, és sok mindent megtettek azért, hogy elkerüljék, például úgy, hogy minden új leletet bűnügyi színhelyként kezeltek, és olyan tisztaszobás technikákat alkalmaznak, amelyek arra vannak tervezve, hogy a csontokat kezelő emberektől származó szennyeződést minél jobban lecsökkentsék.60
Mit tanultunk ebből? Meglepő lenne, hogy a genetika ismét megviccelte az evolucionistákat? A fenti dolgokat figyelmesen megvizsgálva, a neandervölgyi genetikát vizsgáló tanulmányok nagyon meglepő eredményekre jutottak. A korai munkák azt mutatták, hogy rendelkeztek azzal a génnel (FoxP2), ami a mai embernek a beszédképességét adta.61 Utána felfedezték, hogy néhány neandervölgyi olyan bőr-pigment génekkel rendelkezett, ami a világos bőrszínt és vörös hajat okozza, vagy a zöld szemszínért és szeplőkért felel, ha európai származású emberekben fordulnak elő.62
Minden jel arra mutatott, hogy jobban hasonlítanak a mai emberre, mint előtte gondolták, de a neandervölgyi csontokból kivont DNS minták alapján azt a következtetést vonták le, hogy valószínűleg nem keveredtek a mai ember őseivel, mivel a neandervölgyi mitokondriális DNS szekvenciáját nem lehet a mai emberekben megtalálni.63
Ezek a következtetések rövid életűek voltak, mivel az egész neandervölgyi genomot (vagyis annak 60%-át) röviddel utána leközölték.64 Ha a szekvenciák pontosak, akkor a neandervölgyi nem hasonlít semmire, amit ismernénk. Van bizonyíték, miszerint a mai ember közvetlen elődjeivel kereszteződtek, ami a biológiai fajfogalom szerint azt jelenti, hogy ugyanaz a faj vagyunk. Úgy tűnik, hogy az Afrikán kívül élő emberek genetikai anyagának 3-4%-a a neandervölgyiektől származik. Érdekes módon, neandervölgyi maradványokat még nem találtak Afrikában, pedig a genomjuk maradványai fennmaradtak azokon a területeken, ahol éltek (és azon túl is). Az új evolúciós magyarázat szerint az afrikai eredetmodellt úgy módosították, hogy korlátozott keveredés történt a mai ember és a neandervölgyiek között, amint elhagyták Afrikát, és átvették a neandervölgyiek helyét a világuralom felé tartó útjukon. De ez nagy különbség a nem sokkal korábbi elmélethez képest. Emlékszünk még, hogy az evolucionisták mennyire magabiztosak voltak a dolgukban, hogy fejbe verték a bibliai teremtéshívőket az afrikai eredetmodelljükkel? Amit akkor hittek, abból ma a tudományos eredmények sokat cáfolnak.65
Egy másik elképzelés jobban összefér az adatokkal. Ahelyett, hogy két nagy populációval számolunk, amely alig keveredett egymással, lehetséges, hogy a neandervölgyi csoport sokkal kisebb volt, mint a fő migrációs hullám, így teljes keveredés történhetett ugyanazzal az eredménnyel. Ha a modern nem-afrikai populáció pusztán 3-4%-a neandervölgyi, akkor talán a populációjuk csak 3-4%-a volt a teljes méretének, vagy valahol ekörül, különböző fokú keveredés mellett. Ha valódi emberek voltak, különösen, ha az özönvíz után éltek, a keveredés természetes, ahogy teszik ezt az emberek ma is. Újabb kutatások szerint a neandervölgyiek jobban hasonlítottak egymásra, mint bármely modern embercsoport egyedei.66 Spanyolországtól Oroszországig a neandervölgyiek egy kiterjedt emberi családra hasonlítanak – egy emberi családra – ami Európában és Ázsiában élt az özönvíz után, és akiket egy későbbi emberi vándorlás legyőzött.
Ember vagy csimpánz
Évtizedekig azt hallottuk, hogy „az emberek és a csimpánzok 99%-ban azonosak”. Ez nem igaz.67 Ez az adat egyes korai kísérleteken alapul, amelyek egyes ismert gének bizonyos szakaszait hasonlították össze úgy, hogy a DNS-ünknek egy bizonyos része tényleg nagyon hasonlít. Azonban a genomunk kevesebb, mint 2%-a kódol fehérjét, és sok gént nem lehet a két faj között összehasonlítani. Az embernek több száz fehérjét kódoló génje van (mindegyikük szorosan a spliceoszómába integrálva), amelyek a csimpánzból hiányoznak. Egész géncsaládok vannak az emberben, amelyek nincsenek a csimpánzokban.68 Ezzel porszem került az evolúciós modellekbe, mivel elvileg pusztán néhány százezer generáció volt a közös ősünk óta.69 Hogyan keletkezhettek ezek az új gének és kerülhettek be a komplex genomunkba ilyen rövid idő alatt? Az idő nem döntő tényező az evolúciónál. Az evolúciót generációkban mérik, és nem volt elég generáció a feltételezett közös ősünk óta.
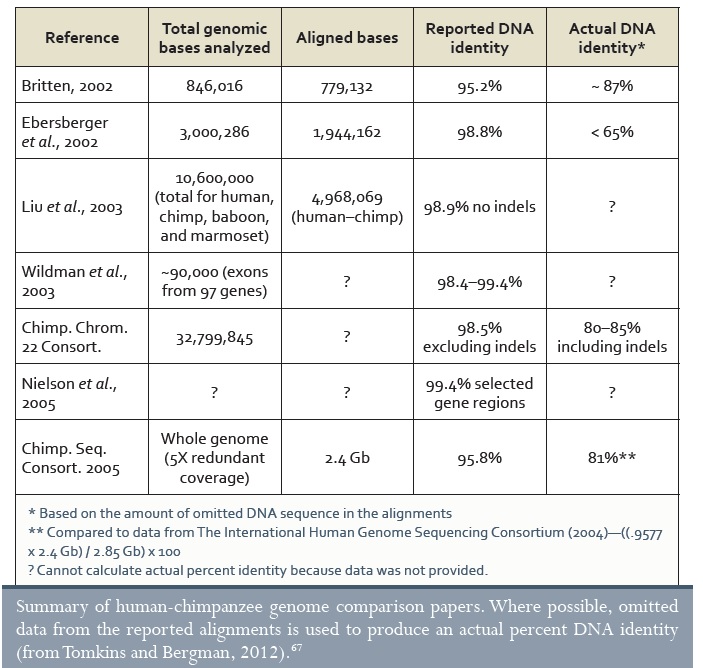
Sőt, mintegy 35.000.000 egybetűs különbség van70, amelyeknek ki kellett alakulniuk (fele-fele arányban mindkét fajban), majd elterjedniük a megfelelő populációkban, végül rögzülniük (az eredeti betű azon a helyen teljesen elveszett) ez alatt a néhány generáció alatt. Hasonlóképpen több tízezer kromoszomális átrendeződésnek kellett végbemennie, elterjednie, majd rögzülnie, több tíz millió bázispár inszerciója (beépülés) és deléciója (elvesztés) mellett. A rögzülések nagyon lassan mennek végbe, és a legtöbb új variáns elvész (mivel definíció szerint ritkák). A mutáció és szelekció sebessége, amely ekkora mértékű változásokhoz szükséges, egy mindössze 6 millió éves időintervallumra vetítve elképesztő, de mégis elengedhetetlen ahhoz, hogy az evolúció igaz legyen.
Az emberi és a csimpánz Y kromoszómák közti hasonlóság hiányáról fent már beszéltünk. A csimpánz Y kromoszóma felénél kimutatott 70%-os azonosság mellett a mutációs ráta bizonyára rendkívül magas. Elgondolkodhatunk azon, hogy miért hasonlítanak egymásra a férfiak Y kromoszómái. Talán azért, mert a kromoszóma mindössze 6.000 éves!
Az ember és a csimpánz esetében számítanak a hasonlóságra. Úgy néznek ki, mint mi, hasonlóan viselkednek, hasonló dolgokat esznek, és hasonló környezetet igényelnek (kivéve, hogy mi voltunk annyira értelmesek, hogy meleg házakat és ruhákat találjunk fel). Miért gondolná bárki, hogy másmilyenek lennénk genetikailag? Gondolkodjunk el azon, hogy mit mondanának az evolucionisták, ha nem volnának hasonlóságok. Azt mondanák, hogy nincs bizonyíték egy értelmes Teremtőre, mivel mindenki azt várná, hogy a Teremtő közös tervezési sablonokat használna hasonló élőlények teremtésére. Ha fej, ők nyernek, ha írás, akkor mi veszítünk!71
Mennyire vagyunk hasonlók? A híres genetikus, Svante Pääbo szavai szerint:
Nincs pontos módja a kiszámolásához … a végén pusztán politikai, társadalmi, illetve kulturális dolog, hogy mi hogyan nézzük a különbségeinket.72
A mutációs ráták túl magasak
Az első fejezetben a magas mutációs ráták hatásairól írtam, valamint arról, hogy a természetes szelekció nem „látja” a legtöbb mutációt. A végkövetkeztetés az volt, hogy a legtöbb mutáció, noha káros, átmegy a szelekciós szűrőn. Ez azt jelenti, hogy a delécióval járó mutációk fajunkban felhalmozódnak, ami ellentétes azzal, amit a hosszú távú evolúció elvár. Ha ezt a tudást a genom komplexitásáról alkotott modern ismereteinkre alkalmazzuk, látható a probléma valódi nagysága. Az eukarióta (sőt minden baktériumon kívüli) genom túl komplex és az eukarióta gének, mint mutációs célpontok túlságosan nagyok ahhoz, hogy az evolúció végbemenjen. Egy átlagos DNS bázis átlagosan hat különböző RNS transzkriptumra íródik át, és több átfedésben lévő kódban használódik fel (hiszton, splicing, fehérje kód, stb.). A mutációk evolucionista szemmel irtózatos mennyiségben fordulnak elő, ami az egész emberiség hosszú távú fennmaradásának szab határt. Egy dolog azonban biztos, mivel Jézus megígérte, hogy visszajön, és azt is megígérte, hogy amikor újra eljön, lesznek még emberek. Emiatt nem fogunk a második eljövetel előtt kihalni. Az mikor lesz? A genomra gyakorolt mutációk káros távlati hatásai miatt ez biztosan nem lesz több millió év múlva.
A kihívás
A genom egy többdimenziós operációs rendszer beépített hibajavító és önváltoztató kódokkal. Van több egymással átfedésben lévő DNS, RNS, illetve szerkezeti kód. Vannak DNS alapú, illetve RNS alapú gének. A genom nagymértékű redundanciával készült, szándékosan. A redundancia ellenére rendkívül tömör, hiszen a mintegy 20.000 „gén” jóval több, mint 100.00 különálló fehérjét tud létrehozni. A genom emellett lassacskán szétbomlik az idők során, holott mindmáig életképes maradt a csodásan megtervezett önkorrigáló kódok és a beépített, intelligens módon megtervezett redundancia miatt.
Darwin híres mondását idézzük:
Ha bizonyítani lehetne, hogy létezik olyan komplex szerv, amely nem alakulhatott ki számos egymást követő csekély módosulás révén, elméletem azonnal összeomlana.73
Tudom, hogy ezt az idézetet sokan kifacsarták (mindkét oldalon) az évek során, de ettől függetlenül állítom, hogy az emberi genom pont egy ilyen dolog. Nem hiszem, hogy pusztán természeti folyamatok során keletkezhetett. Kihívom az evolucionistát, hogy mutasson be egy életképes modellt a genom történetére, ami magába foglalja az információs változások forrását, a szükséges mutációk számát, és leírja a szükséges szelekciós erőket, mindezt a megfelelő időkeretben. Valóban, kihívom őket arra, hogy mondják meg, hogy a genom, bármely genom is, hogyan jelent meg a semmiből egy olyan világban, ahol nem létezett utasítást adó információ.
Ez hova vezet?
Bizonyára látszik, hogy a genetika nem Darwin barátja. Mivel nem ismerte az élet komplexitását, a fajok szaporodásának mikéntjét, a komplex rendszerek törékeny voltát, így tudott egy olyan elméletet szőni, amivel kikerülte ezeket a leküzdhetetlen akadályokat. A darwinizmust a modern ismeretek fényében újra kellene értékelni. A modern genetika viszont a bibliai leírást jól alátámasztja. A modern ember génjeiben számos bizonyíték van két ember (Ádám és Éva) teremtésére, majd a néhány ezer évvel későbbi populációs beszűkülésre (Noé és az özönvíz), valamint az emberi populáció felosztására néhány generációval később (Bábel), amit az emberiség egyszeri, egész világra történő szétterjedése követett. De nemcsak erre van bizonyíték, hanem a mutációs ráták, a mutációk eloszlása, valamint az emberi genomnak nevezett szuper-komplex számítógépes operációs rendszer törékenysége, mind arra mutatnak, hogy ez a rendszer igen fiatal. Azon elgondolkodhatunk, hogy Darwin ugyanarra a következtetésre jutott volna-e, ha ma is élne (feltéve, ha nyitottan és becsületesen kezelte volna ezeket az eredményeket). De nemcsak a genetika létezése jelent veszélyt az evolúcióra, mivel az élet eredete sem barátja az evolúciós elméletnek. A következő fejezetben ebbe nyerünk betekintést, mivel az élet eredetéről alkotott minden evolucionista elképzelés megszegi az ismert kémiai, fizikai, illetve valószínűségszámításra vonatkozó törvényeket.
Hivatkozások:
1. Darwin, C.R., The variation of animals and plants under domestication, London, John Murray, 1868; darwin-online.org.uk.
2. Carter, R., Darwin’s Lamarckism vindicated? March 2011; creation.com/epigenetics-and-darwin.
3. Az angol változathoz, lásd Druery, C.T. and Bateson W., Experiments in plant hybridization, Journal of the Royal Horticultural Society 26:1–32, 1901.
4. Az édesapja elküldte Cambridge-be, miután kudarcot vallott az orvosi tanulmányaiban Edinburgh-ben. Miután a Cambridge-i egyetemen lediplomázott, a híres öt éves útjára indult a világ körül, a Beagle nevű hajón, és sosem lett lelkipásztor.
5. Mendel azt mutatta meg, hogy amikor két egyedet megkereszteznek, amely domináns illetve recesszív génváltozatot hordoznak (pl. domináns lila színű szirmok, illetve recesszív fehér színű szirmok), egy 3:1-es arányt kapunk a vad típus (lila) és a recesszív típus között (fehér). Ez azért történik, mert csak azoknak a növényeknek lesz fehér virágai, amelyek a recesszív génből két példányt tartalmaznak. Ezért a növények három negyede lila virággal rendelkeznek majd, és a negyede pedig fehér virággal rendelkezik majd: ez egy 3:1-es arány a lila és a fehér között.
6. Howard, J.C., Why didn’t Darwin discover Mendel’s laws? JouRNSl of Biology 8:15, 2009.
7. Ide tartozik: Darwin, C.R., The variation of animals and plants under domestication, London, John Murray, 1868; Self-Fertilisation in the Vegetable Kingdom, 1876; and The Different Forms of Flowers on Plants of the Same Species, 1877.
8. Howard, Why didn’t Darwin discover Mendel’s laws?
9. Lodish, H. et al., Molecular Biology of the Cell, 5th ed., W.H. Freeman and Company, New York, 2004.
10. Batten, D., könyváttekintés: The Biotic Message: Evolution versus Message Theory, J. Creation 11(3):292–298; creation.com/biotic.
11. Gitt, W., Scientific laws of information and their implications-part 1, J. Creation 23(2):96–102; creation.com/laws-of-information-1. Lásd ennek a sorozatnak a többi részét is.
12. Lynch, M., Rate, molecular spectrum, and consequences of human mutation, Proc. Nat. Acad. Sci. USA 107(3):961–968, 2010. Hasonló eredményekre jutottak más tanulmányok, mint pl. Neel, J.V. et al, The rate with which spontaneous mutation alters the electrophoretic mobility of polypeptides, Proc. Nat. Acad. Sci. USA 83:389–393, 1986; Nachman, M.W. and Crowell, S.L., Estimate of the mutation rate per nucleotide in humans, Genetics 156:297–304, 2000; Kondrashov, S., Direct estimates of human per nucleotide mutation rates at 20 loci causing Mendelian disease, Human Mutation 21:12–27, 2002.
13. Eyre-Walker, A. and Keightley, P.D., High genomic deleterious mutation rates in hominids, Nature 397:344–347, 1999.
14. Lynch, M., Rate, molecular spectrum, and consequences of human mutation, Proc. Nat. Acad. Sci. USA 107(3):961–968, 2010.
15. Crow, J.F., The origins, patterns, and implications of human spontaneous mutation, Nature Reviews: Genetics 1:40–47.
16. Conrad, D.F. et al., Variation in genome-wide mutation rates within and between human families, Nature Genetics 43:712–714, 2011.
17. Kondrashov, A.S., Contamination of the genome by very slightly deleterious mutations: why have we not died 100 times over, J. Theor. Biol. 175:583–594.
18. ReMine, W.J., Cost theory and the cost of substitution-a clarification, J. Creation 19(1):113–125, 2005; creation.com/cost.
19. Haldane, J.B.S., The cost of natural selection, JouRNSl of Genetics 55:511–524, 1957.
20. Batten, D., Haldane’s dilemma has not been solved, J. Creation 19(1):20–21, 2005; creation.com/haldane. Lásd még saintpaulscience.com/Haldane.htm.
21. Kimura, M., Evolution rate at the molecular level, Nature 217:624–626, 1968.
22. ReMine, Cost theory and the cost of substitution.
23. Ohno, S., So much „junk” DNS in our genome, Evolution of genetic systems, Brookhaven biológiai szimpózium 23. szám (Smith, H.H., ed.), pp. 366–370, 1972.
24. Carter, R.W., The slow, painful death of junk DNS, J. Creation 23(3):12–13, 2009; creation.com/junkDNSdeath.
25. Birney, E. et. al., Identification and analysis of functional elements in 1% of the human genome by the ENCODE pilot project, Nature 447:799–816, 2007.
26. J.S. Mattick, as quoted in Gibbs, W.W., The Unseen genome: gems amid the junk, Scientific American, 47–53. o., Nov 2003.
27. Yan, K.-K. et al., Comparing genomes to computer operating systems in terms of the topology and evolution of their regulatory control networks, Proc. Natl. Acad. Sci. USA 107(20):9186–9191, 2010.
28. Stein, L.D., Human genome: end of the beginning, Nature 431:915–916, 2004.
29. Birney, Identification and analysis of functional elements in 1% of the human genome; lásd még Williams, A., Astonishing DNS complexity update, July 2007, creation.com/DNSupdate.
30. Barash, Y. et al., Deciphering the splicing code, Nature 465:53–59, 2010.
31. Lásd Anon., Human genes sing different tunes in different tissues, PhysOrg.com, 2 Nov 2008.
32. Carter, R.W., Splicing and dicing the human genome, July 2010; creation.com/splicing.
33. Lynch, Rate, molecular spectrum, and consequences of human mutation.
34. Gerstein, M.B. et al., What is a gene, post-ENCODE? History and updated definition, Genome Research 17:669–681, 2007.
35. Schoenfelder, S. et al., Preferential associations between co-regulated genes reveal a transcriptional interactome in erythroid cells, Nature Genetics 42:53–61, 2009; Lásd még: Scientists’ 3-D view of genes-at-work is paradigm shift in genetics, sciencedaily.com, 16 Dec 2009.
36. Eitan, Y. and Tanay, A., Probabilistic modeling of Hi-C contact maps eliminates systematic biases to characterize global chromosomal architecture, Nature Genetics 43(11):1059–1067, 2011.
37. Is the shape of a genome as important as its content? PhysOrg.com, 29 Oct 2010.
38. Bolzer, A. et al., Three-dimensional maps of all chromosomes in human male fibroblast nuclei and prometaphase rosettes, PLoS Biol 3(5):e157, 2005.
39. Lásd: A new look at how genes unfold to enable their expression, PhysOrg.com, 14 July 2008.
40. Baillie, J.K. et al., Somatic retrotransposition alters the genetic landscape of the human brain, Nature 479(7374):534–537.
41. Li, G.-W., Oh, E., and Weissman, J.S., The anti-Shine-Dalgarno sequence drives translational pausing and codon choice in bacteria, Nature 484(7395):538–541, 2012.
42. Például a kétbites rendszerben legalább 5 betűre lenne szükség 20 aminosav kódolására, kevesebb degenerációval (25 = 32 lehetőség).
43. Carter, R.W., The non-mythical Adam and Eve: refuting errors by Francis Collins and BioLogos, August 2011; creation.com/biologos-adam.
44. Sarfati, J., The IncaRNStion: Why did God become Man? December 2010; creation.com/incaRNStion.
45. Carter, The non-mythical Adam and Eve.
46. Sarfati, J., Blood types and their origin, J. Creation 11(1):31–32, 1997; creation.com/blood-groups.
47. Carter, R.W., The Neutral Model of evolution and recent African origins, J. Creation 23(1):70–77; creation.com/african-origins.
48. Cann, R.L., Stoneking, M., and Wilson, A.C., Mitochondrial DNS and human evolution, Nature 325:31–36, 1987.
49. Wieland, C., A shrinking date for eve, J. Creation 12(1):1–3, 1998; creation.com/eve.
50. Madrigal, L. et al., High mitochondrial mutation rates estimated from deep-rooting Costa Rican pedigrees, American JouRNSl of Physical Anthropology 148:327–333, 2012. Lásd még Carter, R.W., Is „mitochondrial Eve” consistent with the biblical Eve? Jan 2013; creation.com/mteve-biblical-eve.
51. Carter, R.W., Mitochondrial diversity within modern human populations, Nucl. Acids Res. 35(9):3039–3045, 2007.
52. Carter, The non-mythical Adam and Eve; Carter, The Neutral Model of evolution and recent African origins.
53. Hughes, J.F. et al., Chimpanzee and human Y chromosomes are remarkably divergent in structure and gene content, Nature 463:536–539.
54. The InteRNStional HapMap 3 Consortium, Integrating common and rare genetic variation in diverse human populations, Nature 467:52–58, 2010.
55. Parvanov, E.D. et al., PRDM9 controls activation of mammalian recombination hotspots, Science 327:835, 2010; Berg, I.L. et al., PRDM9 variation strongly influences recombination hot-spot activity and meiotic instability in humans, Nature Genetics 42(10):859–864, 2010; Lásd még, Carter, R.W., Does genetics point to a single primal couple? Apr 2011; creation.com/genetics-primal-couple.
56. Tishkoff, S.A. et al., The genetic structure and history of Africans and African Americans, Science 324:1035–1044, 2009.
57. Carter, The Neutral Model of evolution and recent African origins.
58. Behar, D.M. et al., and The Genographic Consortium, The dawn of human matrilineal diversity, Am. J. Human Gen. 82:1130–1140, 2008.
59. Carter, R.W., The Painted Neanderthal, May 2010; creation.com/the-painted-neandertal.
60. Carter, R.W., Neandertal genome like ours, June 2010; creation.com/neandergenes.
61. Borger, P. and Truman, R., he FOXP2 gene supports Neandertals being fully human, J. Creation 22(2):13–14; creation.com/foxp2.
62. Lalueza-Fox, C. et al., A Melanocortin 1 Receptor allele suggests varying pigmentation among Neanderthals, Science 318:1453–1455, 2007.
63. Green, R.E. et al., A complete Neandertal mitochondrial genome sequence determined by high- throughput sequencing, Cell 134:416–426, 2008; Carter, R.W., The Neandertal mitochondrial genome does not support evolution, J. Creation 23(1):40–43, 2009; creation.com/neandertal-mito.
64. Green, R.E. et al., A draft sequence of the Neandertal genome, Science 328:710–722, 2010.
65. Carter, R.W., Neandertal genome like ours (There may be Neandertals at your next family reunion!), 2010 június; creation.com/neandergenes.
66. Reich, D. et al., Genetic history of an archaic hominin group from Denisova Cave in Siberia, Nature 468:1053–1060, 2010. Lásd még Wieland, C. and Carter, R.W., Not the Flintstones -it’s the Denisovans, Jan 2011; creation.com/mteve-biblical-eve.
67. Tomkins, J. and Bergman, J., Genomic monkey business—estimates of nearly identical human-chimp DNS similarity re-evaluated using omitted data, J. Creation 26:94–100, 2012; creation.com/human-chimp-DNS-similarity-re-evaluated.
68. Demuth J.P. et al., The evolution of mammalian gene families, PLoS ONE 1(1): e85, 2006.
69. 6 millió év / ~20 év generációnként = pusztán 300.000 generáció.
70. Varki, A. and Altheide, T.K., Comparing the human and chimpanzee genomes: searching for needles in a haystack, Genome Research 15:1746–1758, 2005.
71. Statham, D., Heads I win, tails you lose: the power of the paradigm, Nov 2010; creation.com/fused.
72. Cohen, J., Relative differences: the myth of 1%, Science 316:1836, 2007.
73. Darwin, C., Origin of Species, 6. kiad., 1872.