EAH 3. Élet eredete
Részlet az Evolúció Akhilleusz sarkai (halálos pontjai) című könyvből

3. fejezet:
Az élet eredete
78-111 oldal
A könyvet Robert Carter, Ph.d szerkesztette.
Kiadó: Creation Book Publishers. Második kiadás, 2014. október
Honlap: Creation.com
Fordította: Dr. Nagy Zoltán növénybiológus.
Lektorálta: Dr. Hecker Kristóf biológus.
Dr Don Batten Ph.D: Az evolúció Akhilleusz sarkai című könyvének fejezetenkénti fordítása.
Tartalomjegyzék:
1. fejezet: Természetes szelekció
6. fejezet: Radiometrikus kormeghatározás
8. fejezet: Etika és erkölcsiség
3. fejezet
Dr Jonathan Sarfati
Fizikai kémia, Ph.D.
Victoria University of Wellington, Új-Zéland
Dr. Sarfati közismert a teremtéstudománnyal foglalkozó körökben: számos publikáció és négy önálló könyv megírása kötődik a nevéhez és természetesen sok könyv társszerzője is. A „Refuting Evolution” (Megcáfolt evolúció) című könyve a valaha legsikeresebb kreacionista könyv a világon. Munkáiban éles és logikus gondolkodása tükröződik vissza, talán éppen sakkozó múltjából eredően (F.I.D.E. Master (FM) címet nyert 1988-ban). Jonathan kapta azt a feladatot, hogy írjon a darwini evolúció harmadik gyenge pontjáról, az élet eredetének kérdéséről. Ebben a fejezetben egyértelműen és ésszerűen világít rá arra a leküzdhetetlen problémára, amit az élettelen vegyületekből kialakuló első élőlények jelentenek.
Nézze meg honlapunkat: creation.com/dr-jonathan-sarfati
Bevezetés
Az előző fejezetben a genetika által felvetett problémákkal foglalkoztunk. Bár azok a bizonyítékok igen nagyvonalúak voltak az evolúciós elmélettel, mert feltételeztek egy működő genetikai rendszerrel bíró önreprodukáló sejtet. Egy még korábbi fejezetben pedig, ahol a természetes szelekcióval foglalkoztunk, hasonlóan nagyvonalú volt az érvelés, hiszen a felsorolt tények olyan esetben működnek, ha léteznek olyan rendszerek, melyek képesek bármilyen kiválogatott információt továbbadni. Ez a fejezet pedig megadja a korábbi cáfolatok alapját. Azt fogjuk látni, hogy nagyon sok minden szükséges az első, önmagát szaporítani képes rendszer kialakulásához.
Kiderül majd, hogy az első élet keletkezése a leggyengébb pontja a materialisták elméletének. Még a legegyszerűbb egysejtű élőlények is extrém komplex felépítésűek, számos, bonyolult „gép” található bennük, valamint a szintézisükhöz szükséges információ is bennük tárolódik, és ezt mind képes a sejt kiolvasni és az utódokba átörökíteni. A legegyszerűbb elméleti szaporodóképes élőlénynek is számos fehérjére és molekuláris mechanizmusra van szüksége, hogy kódolni és tárolni tudja azt az információt, ami a működéséhez elengedhetetlen.1
Lehetséges, hogy egy ilyen sejt egyszerű vegyületekből önmagától kialakuljon?
Minden jelenleg ismert, magától szaporodni képes élőlény a Földön DNS-ben tárolja a biológiai információt, de ez az információ nem értelmezhető egy dekódoló rendszer nélkül. A dekódoló rendszer elemeinek az információja is a DNS-ben van tárolva és kódolva. Meg lehet oldani ezt a nehéz „tyúk vagy tojás” problémát? Továbbá a legtöbb biológiai folyamat energiát használ, ami ATP-ben tárolódik, amit egy ATP szintetáz nano-motor állít elő. De az ATP szintetáz motor létrejöttéhez is a DNS-ben tárolt utasításkészlet szükséges, amit szintén az ATP-t használó olvasó rendszer dolgoz fel – ez már egy három körös ciklus, ami már egy „pete-nimfa-szöcske” probléma (ami sokkal rosszabb, mint egy „tyúk vagy tojás” probléma). Van megoldása ennek a zavarba ejtő rejtélynek, vagy az élet keletkezése lehetetlen úgy, ahogy mi azt elképzeljük?
Néhányan úgy próbálják megoldani ezeket a problémákat, hogy feltételeznek egy olyan molekulatípust, ami egyedül képes az információ tárolásra/dekódolásra. Erre a pozícióra a legtöbben az RNS-t tartják alkalmasnak, de tényleg elegendő ez az első élet kialakulásához?
Továbbá, a feltételezett őslevesben a szabályozatlan kémiai folyamatok nagyon kis valószínűséggel képesek csak komplex vegyületeket létrehozni. A biológiai építőkövekből (pl. aminosavak) megfelelő hosszúságú molekuláknak kell összeállnia az élethez; azonban ezek a hosszú molekulák instabilak. Az építőkövek szerkezete sem mindegy, hogy milyen, illetve a hígulás és a szennyeződések sem hasznosak; az őslevesben „bal- és jobbkezes” vegyületek egyforma arányban jönnek létre, de az élőlényekben csak balkezes formák fordulnak elő. Ezek a tények átírják az „élet eredetének” a forgatókönyvét?
Végül, a kémiai evolúció kutatásainál, a kémcsöves és számítógépes szimulációk esetén olyan elfogadhatatlan hatásokat visz be a rendszerbe egy intelligens kutató, ami nem lehetett jelen a feltételezett őslevesben. Ezek ily módon szolgáltathatnak elegendő bizonyítékot az élet spontán keletkezése mellett?
Mindezek tetejébe a vezető kutatók elismerik, hogy a kémiai evolúciót nem a tények és bizonyítékok alapján fogadják el, hanem materialista hitük miatt. Miért van az mégis, hogy ezt tényként állítják szembe a Biblia állításával, ami szerint az intelligens Isten hozta létre az életet?
A darwini folyamatok nem tudják magyarázni az élet keletkezését
 Mindig is volt feszültség a kémiai folyamatok eredménye által létrejött élet elmélete és a darwini evolúció között, de a legtöbb ember nem tud erről az ellentmondásról. Látszólag maga Darwin is komoly akadályokat látott abban, hogy az élet keletkezése hogy illeszkedik az elméletébe. Számos észrevételt tett nyomtatásban is és még több megjegyzést a személyes levelezéseiben, de legtöbb írásában próbálja elkerülni a konklúzió levonását. Ismerte azokat a régi elméleteket, amik az élőlények spontán szaporodást cáfolták (mint például egerek vagy legyek esetében), ilyen volt Francesco Redi elmélete 1668-ból, de még erősebb érveket sorakoztatott fel a kortárs kreacionista tudós, Louis Pasteur, aki 1861-ben2, két évvel a „Fajok eredete” megjelenése után, kísérleteivel teljesen leleplezte spontán keletkezés lehetőségét – még a mikróbák esetében.
Mindig is volt feszültség a kémiai folyamatok eredménye által létrejött élet elmélete és a darwini evolúció között, de a legtöbb ember nem tud erről az ellentmondásról. Látszólag maga Darwin is komoly akadályokat látott abban, hogy az élet keletkezése hogy illeszkedik az elméletébe. Számos észrevételt tett nyomtatásban is és még több megjegyzést a személyes levelezéseiben, de legtöbb írásában próbálja elkerülni a konklúzió levonását. Ismerte azokat a régi elméleteket, amik az élőlények spontán szaporodást cáfolták (mint például egerek vagy legyek esetében), ilyen volt Francesco Redi elmélete 1668-ból, de még erősebb érveket sorakoztatott fel a kortárs kreacionista tudós, Louis Pasteur, aki 1861-ben2, két évvel a „Fajok eredete” megjelenése után, kísérleteivel teljesen leleplezte spontán keletkezés lehetőségét – még a mikróbák esetében.
A Fajok eredetének első kiadásának (1859) utolsó fejezetében Darwin ezt írta:
Az analógiákból levezethető, hogy valószínűleg minden, a Földön valaha élt szerves lény ugyanattól az egynéhány ősi formától származik, amibe először lehelték az életet.3
Ez alig tekinthető az élet spontán létrejöttét támogató kijelentésnek. Amint azt egy barátjának 1863-ban írt levelében megvallja, megbánta, hogy nem egyértelműen fejezte ki magát ebben a témában:
De megbántam, hogy megalázkodtam a nyilvánosság véleménye előtt, és a bibliai teremtés kifejezést használtam, holott inkább a ’megjelenik’ kifejezésre gondoltam, ami egy teljesen ismeretlen folyamat … Ez mai ismereteink alapján egy meglehetősen ostoba gondolat az élet létrejöttéről; ugyanígy gondolhatnánk az anyag eredetére.4
Mégis, furcsa módon, ezt az utolsó mondatot szúrta be a Fajok eredetének minden későbbi kiadásába:
Felemelő elképzelés ez, amely szerint a Teremtő az életet a maga különféle erőivel együtt eredetileg csupán néhány, vagy csak egyetlen formába lehelte bele, mialatt bolygónk a gravitáció megmásíthatatlan törvényét követve keringett körbe-körbe, és ebből az egyszerű kezdetből kiindulva végtelenül sokféle, csodálatos és gyönyörű formák bontakoztak ki – és teszik ezt ma is.
Teljesen világos hogy Darwin hitt az evolúcióban, de nem világos, hogy a kémiai evolúcióban is hitt-e? 1871-ben (ebben az az évben jelentette meg „Az ember származása” c. könyvét, amiben az ember alacsonyabb szintű létformákból való kifejlődéséről ír) komoly ellentmondásba került:
…ha (és óh, micsoda nagy ha) elfogadnánk egy kis meleg pocsolyát az ammónia és foszfor sóféleségeivel, fény, meleg és elektromosság jelenlétében, akkor egy kémiai úton keletkezett fehérje azonnal további bonyolult változásokon mehetne keresztül…5
Tehát még Darwin is, a modern evolúcióelmélet atyja is két lehetőség között vívódott a legfontosabb témában – hogy hogyan keletkezhetett az élet?
A híres filozófus, Antony Flew (1923-2010) is rámutatott erre a problémára, és közvetlenül így fejezte ki aggályait a vezető ateista evolucionistának, Richard Dawkins-nak6 (1941-):
Úgy tűnik, hogy Richard Dawkins folyamatosan figyelmen kívül hagyja azt a tényt, hogy Darwin a Fajok eredetének tizennegyedik fejezetében az egész érvelését arra építi fel, hogy léteznie kell egy szaporodási képességgel rendelkező élőlénynek. De egy valóban hihető elméletnek egy ilyen élőlény kialakulására is feltétlenül magyarázatot kell adni.
Darwin azonban jól tudta, hogy nem adott erre magyarázatot. És úgy tűnik számomra, hogy több mint ötven évvel a DNS felfedezése és az ismert kutatási eredmények alapján új, megbízható és erős bizonyítékok állnak a tervezettség mellett.7
Meg kell jegyezni, hogy Dr. Flew egészen mostanáig az ateizmus kiemelkedő támogatójaként volt ismert, de halála előtt az ateista közösség legnagyobb megdöbbenésére elvetette ezt a hitét.8 A fő érv ebben a döntésében a legegyszerűbb önreprodukáló sejt elképesztő bonyolult felépítése volt.
Theodosius Dobzhansky (1900-1975), a 20. század egyik vezető evolucionistája és buzgó materialista (orosz ortodox neveltetése ellenére9) szintén hasonló eredményre jutott. A Fajok eredetének kommentárjában határozottan elutasítja azokat az elméleteket, amik a természetes szelekcióval érvelnek, mert azok feltételeznek egy már létező élőlényt:
Néhány, az élet eredetéről szóló irodalmat olvasva, sajnos azt látom, hogy a szerzők rosszul használják ezt a kifejezést [természetes szelekció]. A természetes szelekció az élőlény differenciált szaporodásáról és a tulajdonságok öröklődéséről szól. A természetes szelekció rendszerének szükséges feltétele az önreprodukció vagy az élőlény önmaga másolásának a képessége és legalább két ilyen élőlénynek kell lennie. … Könyörögve kérem, hogy ne használjuk a ’természetes szelekció’ szót, ha ezek a feltételek nem adottak. A természetes szelekció prebiológiai környezetben értelmetlen fogalom.10
Érdemes ezt a különbséget a későbbiekben is észben tartani.
Kémiai evolúció
Sok evolucionista próbálja meg elutasítani az ebben a fejezetben felvetett érveket azzal, hogy az evolúciónak nem kell megmagyaráznia az élet létrejöttét élettelen vegyületekből, mivel ezt az abiogenezis írja le. De az általuk elismert evolucionista, Gordy Slack, ezt nem támogatja:
Úgy gondolom álságos dolog úgy érvelni, hogy az élet eredetéhez semmi köze az evolúciónak. Legalább annyira nagy a jelentősége, mint a „Nagy Bumm” esetében a fizikának vagy a kozmológiának. Az evolúciónak meg kell tudnia magyarázni, legalább elméleti szinten, hogy az első szaporodásra képes élőlény hogyan jött létre biológiai vagy kémiai folyamatok által. És ahhoz, hogy megértsük ezt az élőlényt, tudnunk kell mi volt előtte. De ettől nagyon távol vagyunk még.11
A zoológus és fiziológus (és evolucionista) Gerald Kerkut (1927-2004) az evolúció általános teóriáját (ezt az első fejezetben tárgyaltuk) úgy definiálta, mint azt „az elméletet, ami minden földi élőlény keletkezését magyarázza egyetlen ősből, ami maga szervetlen anyagból jött létre.”12 Azonban az evolúciónak ez a része nem darwinista, ahogy ezt már említettük. Ezt kémiai evolúciónak nevezik. Például 1978 szeptemberében az evolúció mellett elkötelezett folyóirat, Scientific American, egy terjedelmes cikket jelentetett meg „Kémiai evolúció és az élet eredete” címmel. Ezt olvashatjuk benne:
J.B.S. Haldane, brit biokémikus13, vetette fel először, hogy redukáló, szabad oxigén nélküli légkör kellett az élet evolúciójához az élettelen, szervetlen anyagból.14
Az élet eredetének elméletével foglalkozó „régi motoros” tudós, Cyril Ponnamperua, a maryland-i egyetem kémiai tanszékén működő Kémiai Evolúciós Laboratórium munkatársa, társszerzője egy hasonló című publikációnak.15 A Discover magazin is kiadott egy különszámot 2011-ben, „Evolúció: az élet történetének újragondolása” címmel, és egy cikk valóban az élet eredetéről szólt.
Az egyszerű sejt?
Darwin idejében sok ember elfogadta a spontán keletkezést – hogy az élet az élettelen anyagból keletkezik. Ezt valahogy könnyű volt akkoriban elhinni, mert a sejt felépítése majdnem teljesen ismeretlen volt. Ernst Haeckel, Darwin német „bulldogja”, azt állította, hogy a sejt csak „szénatomok egyszerű fehérjeszerű keveréke”.16 Ez nem igazán elfogadható nézet, mert már Haeckel előtt a fénymikroszkópnak köszönhetően sok sejtszervecske ismert volt.17
Sőt, a molekuláris biológia forradalma az elmúlt fél évszázadban megmutatta, hogy a sejtnek nem csak magas információtartalomra van szüksége, hanem azt az információt át is kell tudnia örökíteni az utódaiba (reprodukció).
Információ vs kémia
Az előző fejezetben láttuk, hogy a DNS milyen hatalmas mennyiségű kódolt információt tartalmaz. Az ateista Richard Dawkins maga mutat erre rá:
Egyetlen emberi sejt információkapacitása akkora, hogy képes az Encyclopedia Brittanica mind a 30 kötetét háromszor vagy négyszer eltárolni.18
A különbség az élő és élettelen között nem az alkotóelemekben van, hanem az információban. Az élő dolgok hatalmas mennyiségű információt tartalmaznak. Az információ legnagyobb része a DNS-ben van kódolva…19
Dawkins elmagyaráz sok mindent a természetes szelekcióról, de ahogy látjuk, ez nem működik az első élő sejt nélkül. És ennek a magyarázata a kémiára van hárítva. Azonban, a DNS építőelemeinek kémiájában nincs semmi, ami magyarázná azok előre meghatározott kapcsolódási sorrendjét, legalábbis semmivel sem több mint a tintamolekulák közötti erők, amik által a betűk és szavak formálódnak. Polányi Miklós (1891-1976), a Manchester Egyetemen (UK) fizikai kémiai tanszékének egykori vezetője, aki a filozófia felé fordult, ezt állapította meg:
Amint a nyomtatott lapnak az elrendezése (bekezdések, mondatok) független a papír és tinta kémiájától, úgy a DNS molekula szekvenciája is független a DNS molekulában működő kémiai folyamatoktól. A szekvenciának ez a fizikai függetlensége eredményezi azt, hogy bármilyen részleges szekvencia létrejöhet, és ezáltal lesz jelentése – a jelentés egy matematikailag meghatározható és mérhető információtartalom.20
Például: az információ ebben a könyvben nem függ a tintamolekulák tulajdonságaitól a papíron (vagy a pixelekétől a monitoron, ha elektronikus formában olvassa), de mégis betűkké, szavakká, mondatokká és bekezdésekké rendeződnek. Ha a tinta kifolyik, nem képes egy Shakespeare darabot létrehozni! A betűk is értelmetlenek, hacsak nem értjük a nyelvet, ami alapján rendeződnek. Például a ’gift’ szó jelentése angolul ’ajándék’, de német nyelven ’méreg’. A téves megközelítés téves üzenetet eredményez. Egy német barátom mondta nekem, hogy mikor először volt angol nyelvterületen, azt gondolta, hogy őrültek vagyunk, mert mindenhol ’méreg boltok’ vannak.
Az információ az élet kulcsa, nem a véletlen. Az információ nem véletlenszerű, és nem is ismétlődően rendezett, mint a kristályok esetében, amik csak kevés információt tartalmaznak.
Leslie Orgel (1927-2007), egy, az élet eredetét kutató vezető evolúcióbiológus a 20. században, ezt mondta:
Az élőlényeket a specifikus komplexitás teszi különlegessé. A kristályok, mint a gránit, nem tekinthetők élőnek, mert a komplexitás hiányzik belőlük; a véletlenszerűen összeálló polimerekből pedig a specifitás hiányzik.21
Sőt, az információ a jelek nem ismétlődő, nem előrejelezhető elrendeződése, amit egy előzetesen létrehozott rendszer képes olvasni és értelmezni, ill. előre meghatározott szabályok szerint tárolni, javítani és dekódolni. Az információ minden mai formájában intelligens eredetű, hogy az üzenetet továbbítani tudja. Az élet pedig az információn alapszik.
Ahogy azt egy evolucionista fizikus, Paul Davies (1946-) mondta:
Most már tudjuk, hogy az élet titka nem a kémiai összetevőkön múlik, hanem a molekulák logikus szerkezetén és szervezett, megfelelő rend szerint történő elhelyezkedésén. … Mint egy szuperszámítógép, az élet egy információ-feldolgozó rendszer. … Az élő sejtnél az igazi titok a szoftver, nem pedig a hardver.22
De nem tett semmit, hogy megoldja ezt a problémát. Sőt, így folytatja, „Hogyan lehetséges, hogy buta atomok spontán megírják saját programjukat?” Azt válaszolja, „Senki nem tudja…” majd elismeri, „Ismeretlen az a fizikai törvény, ami képes a semmiből információt létrehozni.”
Az információ eredete egy hatalmas gyenge pontja az evolúciós elméletnek. Egyszerűen nincs természetes analógja ennek a jelenségnek. Az információ antitézise a véletlenszerűség. Ugyanígy a túlzott rendezettség (mint a kristályokban). Annak, hogy az információ, ami nem véletlen jelekből áll, nem is végtelenül ismétlődik, mégis magától jöjjön létre a természetben, mindenféle gondolkodó vagy programozott gépezet nélkül, olyan kicsi a valószínűsége, hogy azt jogosan nevezik úgy, hogy „lehetetlen”.
Alex Williams ezt a következőképpen magyarázza el egy működő sejt esetében:
Polányi az élő szervezetekben található gépszerű szerkezetekre mutat … Ahogy a közönséges gépek szerkezete és funkciója nem magyarázható az őket alkotó fémek tulajdonságaival, ugyanígy az élőlények hasonló rendszereinek tulajdonságai sem magyarázhatóak a szén, hidrogén, oxigén, nitrogén, foszfor, kén és más nyomelemek tulajdonságaival. Végtelen számú példa található a tovább nem egyszerűsíthető szerkezetekre az élő rendszerekben, de mindegyik alá van rendelve az ’autopoézis’ [„önszervezés”] alapelvének.23
Egy másik cikkben is erről beszél Williams:
Az autopoézis egy egyedi és fantasztikus tulajdonsága az életnek – nincs ehhez hasonló az ismert univerzumban. Ez az alapja a legalapvetőbb struktúrák szerveződési szintjeinek. Ezek a szintek: (i) az alkotóelemek tökéletes összerendeződése, (ii) az alkotóelemek nagyon specifikus szerkezete, (iii) az alkotóelemek összehangolt működése, (iv) a széleskörűen szabályozott, információvezérelt folyamatok és (v) az ok-okozati meta-információs stratégia az egyedek és fajok túléléséért … mindegyik szint létrejön, de nem magyarázható az alatta lévő szint tulajdonságaival. Az alap szint (alkotóelemek tökéletes összerendeződése) és a természeti környezet között is áthidalhatatlan szakadék van.
Vagyis, a biomolekulák speciális alakja nem következik a hidrogén, oxigén és szén tulajdonságaiból. Ehhez hasonlóan a DNS bázisok sorrendje sem következik az egyes bázisok kémiai tulajdonságaiból. Minden szintnél megtaláljuk ezt az ellentmondást. Williams ezekre az ellentmondásokra Polányi-féle lehetetlenség néven utal és részletezi az élő és élettelen rendszerek közötti szakadékokat. Ezek a szakadékok újabb gyenge pontjai az evolúciós elképzelésnek. Ha azt hinnénk, hogy a természetben van mód ezek áthidalására, az olyan, mintha a világ összes hidrogén atomja hirtelen egy egyenes vonalba rendeződne az univerzum egyik sarkától a másikig, véletlenszerűen. Világos, hogy ez soha nem történik meg, még ha elméletileg lehetséges is. Ez áll a Polányi-féle lehetetlenség elgondolása mögött.
A „tyúk vagy tojás” probléma
nem szűnt meg
Figyelemre méltó, hogy a DNS „nyelvet” lefordító dekódoló rendszert is a DNS kódolja, ez egy igazi „tyúk vagy tojás” probléma. Az ismert tudományfilozófus, Sir Karl Popper (1902–1994) is már rámutatott erre:
Mi teszi az élet eredetét és a genetikai kódot egy igazán zavarba ejtő rejtvénnyé? A genetikai kód nem rendelkezik semmilyen biológiai funkcióval, amíg le nem fordítódik; vagyis amikor szintetizálódnak a fehérjék, melyeknek a szerkezetét kódolja. De … ez a gépezet, amikkel a sejtek … a fordítást végzi, legalább ötven makromolekula összehangolt működését igényli, amelyek szerkezetét szintén a DNS kódolja. A kód nem fordítható le, csakis a fordítás egyes termékeinek felhasználásával.
Ez egy olyan zavarba ejtő kör, egy igazán makacsul feloldhatatlan kör, ami próbára tesz mindenféle elméletet a genetikai kód eredetére vonatkozóan.
Így azzal kell szembesülnünk, hogy az élet eredetének lehetősége (hasonlóan a fizikai törvények eredetéhez) olyan megoldhatatlan akadály a tudományban, hogy nem lehet a biológiát a kémia és fizika szintjére lebontani.24
Habár Popper már négy évtizeddel ezelőtt leírta, hogy a genetikai kód eredetének rejtélye a mai napig nem oldódott meg, Dawkins ezt 2009-ben ismerte el:
Ez az élet eredetének ’22-es csapdája’: A DNS képes másolódni, de szükségesek fehérjék, enzimek, amik katalizálják ezt a folyamatot. Fehérjék képesek a DNS szintézist katalizálni, de szükségük van DNS-re a helyes aminosav sorrend kialakításához.25
Valóban, a genetikai kód eredete egy ördögi kör: fehérjék szükségesek a DNS olvasásához, de e fehérjék felépítését és összehangolt működését is a DNS kódolja. Továbbá, az egész rendszer energiát igényel, ATP formájában, melyet az ATP-szintáz nanomotor állít elő, amit szintén a DNS kódol, amit ATP-t használó gépek dekódolnak! Egy teljesértékű sejt működéséhez egyszerre van szükség a fehérjékre, mint gépekre és a DNS-re, mint információtároló anyagra. És természetesen, az egész haszontalan a szaporodáshoz szükséges információ nélkül.
Csodálatos szerkezetek a DNS dekódolásában
A transzkripció és az átíró gépek
Még a DNS fehérjéket kódoló részének mRNS-re történő átírása is bonyolult gépezetet igényel. Az RNS polimeráz enzim végzi el ezt a feladatot, mely négy fehérjeláncból áll. És egy másik fehérje szükséges ahhoz, hogy utasítsa az RNS polimerázt, hogy hol kezdje el olvasni a DNS mintát. Ezután ez az enzimkomplex mozog a DNS szálon, folyamatosan hozzáadva az illeszkedő RNS betűket, hogy végül pontosan a megfelelő helyen álljon meg.
Richard Ebright és kutatócsoportja a Rutgers Egyetemen további nehézségekkel találkoztak a transzkripció folyamata során.26 Hiszen ez az mRNS átirat fordítódik le fehérjékké a riboszómáknak hívott összetett gépekben.
A DNS kétszálú, de csak egy szál íródik át, tehát a másoláshoz a DNS-t ki kell tekerni. A másoló molekula, az RNS polimeráz (RNSP), először rákapcsolódik a gén (pl. fehérje kódoló DNS szakasz) elejére. A felcsatlakozott RNSP a DNS szálon haladva azt kitekeri.27 Szétnyílik a kettős szál, hogy az mRNS másolat elkészülhessen az egyik oldalon. Ugyanakkor, az összetekert szál energiát tárol; hasonlóan a gumiszalaghoz a gumimotoros játékrepülőben. És ahogy a játékrepülőnél is, az energia végül felszabadul és képes a légcsavart meghajtva a levegőbe emelni a repülőt. Ez vissza is csavarja a kitekert DNS-t (’visszatekeredés’), ami azután elszakad a gépezetről.
Transzláció és a riboszóma
A riboszóma egy olyan molekuláris gépezet a sejtben, ami az mRNS által hordozott információ alapján elkészíti a fehérjéket. Még az ’egyszerű’ E. coli baktériumban is, mely az ember emésztőrendszerében él, a riboszómák 50 különböző fehérjéből és három féle riboszomális RNS-ből (rRNS) épülnek fel, a sejtmaggal rendelkező élőlényekben (eukariótákban) pedig már 73 féle fehérje és 4 féle RNS található. Egy szakértő ezt írta erről a gépezetről:
A riboszóma, a hozzá tartozó kiegészítőkkel, valószínűleg a valaha létezett legbonyolultabb gépezet. Minden alkatrésze aktív és mozog, miközben környezetbarát, és csupán GDP-t és foszfátot termel.28
A riboszóma arról is gondoskodik, hogy a fehérje egyenes láncokat alkosson. A gépen kívül, a növekvő peptid lánc könnyen alakít ki nemkívánatos oldalláncokat, melyek reakcióba léphetnek egymással (egyes aminosavak, pl. az aszpartámsav és a glutaminsav -COOH csoportja képes a lizin és arginin -NH2 oldacsoportjaival reakcióba lépni). Az ipari peptid szintézis esetében ezeket az oldalcsoportokat gátolni kell megfelelő védőcsoportokkal, és mikor a szintézis befejeződött, akkor ezeket el kell távolítani. De a feltételezett őslevesben ezek a feltételek és szerveskémiai megoldások nem voltak elérhetőek. Az élet naturalista eredete tehát komoly problémákkal szembesül a valódi kémiai ismeretek fényében. Az élet alapját olyan molekulák (DNS, RNS és fehérjék) alkotják, amik nem valamilyen külső, egyedi rendeltetésű kémiai rendszerben, vegyi üzemben készültek (beleértve az élő sejteket is).
Nincs élet a riboszómák nélkül. Valóban, az élet nem lenne elképzelhető nélkülük, hiszen kémiailag nem lehetséges a fehérjék ismétlődő előállítása összetett folyamatellenőrzés nélkül, amiről a riboszóma gondoskodik minden élőlényben.
Transzfer RNS
A transzfer RNS (tRNS) molekulák létfontosságú adapterek, melyek szerkezete négylevelű lóheréhez hasonló, és szerepük az egyes aminosavak szállítása az újonnan készülő fehérje szintéziséhez. Körülbelül 80 nukleotidból épülnek fel, melyekből három alkotja az ún. antikodont. Az antikodonnal csatlakozik az mRNS három nukleotidból álló kodon szakaszához. Tehát a tRNS-ek kapcsolják a megfelelő aminosavakat a megfelelő helyre a növekvő peptidlánchoz.
Ezen felül, mindegyik aminosavnak aktivált állapotba kell kerülnie, egy energiaszintet át kell lépnie, hogy képes legyen hozzákapcsolódni a szomszédos aminosavhoz. Az energiát ehhez ATP biztosítja (lásd lentebb). Majd egy speciális enzim, az aminoacil-tRNS-szintetáz (aaRS) köti hozzá az egyes aminosavakat, két lépésben, a megfelelő tRNS-hez. Legalább 20 féle különböző aaRS szükséges, minimum ahány féle aminosav van. Ha itt bármilyen hiba történik, a genetikai kód nem tud megfelelően működni.
A tRNS adapter molekulának pontosan a megfelelő geometriával kell rendelkeznie, hogy
1) az aminosavat megfelelő pozícióban tartsa a peptidlánc létrejöttéhez és
2) hogy az antikodont az mRNS megfelelő szakaszához illessze.
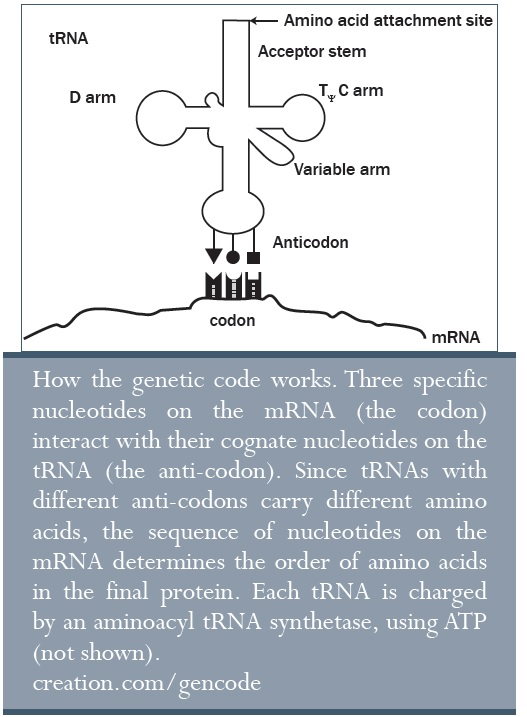 Az adapter molekula pontos geometriájának csakis a riboszómán belül van értelme (tehát vagy együtt fejlődtek ki, ami megoldhatatlan problémát vet fel, vagy egy briliáns Tervező műve mindkettő). Egy bármilyen ősriboszomális élet esetében nincs szükség a speciális geometriára. De akkor honnan jött ez a sajátos szerkezet? Ráadásul, az elméletben kialakuló adapterek valószínűleg egymást, de az mRNS működését zavarják.29 A riboszóma speciális geometriájával ellenőrzött környezetet biztosít a sejt számára, amiben különleges, de elengedhetetlen feladatukat el tudják végezni.
Az adapter molekula pontos geometriájának csakis a riboszómán belül van értelme (tehát vagy együtt fejlődtek ki, ami megoldhatatlan problémát vet fel, vagy egy briliáns Tervező műve mindkettő). Egy bármilyen ősriboszomális élet esetében nincs szükség a speciális geometriára. De akkor honnan jött ez a sajátos szerkezet? Ráadásul, az elméletben kialakuló adapterek valószínűleg egymást, de az mRNS működését zavarják.29 A riboszóma speciális geometriájával ellenőrzött környezetet biztosít a sejt számára, amiben különleges, de elengedhetetlen feladatukat el tudják végezni.
Továbbá, a tRNS adapterekről le kell kapcsolódnia az aminosavaknak miután azok a növekvő fehérjéhez hozzákötődtek. A riboszóma, mint egy kilincs mozog az mRNS-en, és a lekapcsolódáshoz egy másik energiatároló molekulát használ, a GTP-t (guanozin trifoszfát), amit egy összetett, integrált és szigorúan szabályozott gépezet állít elő.30
Annyit érdemes megjegyezni, hogy ez egy hihetetlenül összetett rendszer. Hogyan keletkezhetett volna mindez véletlenszerű kémiai folyamatok révén az őslevesben? Túlságosan sok nehézséggel találkozik a fejlődő rendszer, és az alapvető kémia szemszögéből is túl sok az ellenvetés a folyamattal szemben.
Kettős szűrők: fejlett kémiai gépek
Az aminoacil-tRNS-szintetázok (aaRS-ek) speciális enzimek (fehérje gépek) melyek csodálatos reakciókat hoznak létre. Képesek különbséget tenni két kémiailag nagyon hasonló aminosav között is (pl. a leucin és az izoleucin között, melyeket laborban is különösen nehéz elválasztani).31 De ez a kis különbség is élet és halál kérdése lehet az élettanilag fontos molekulák esetében.
Az izoleucin aaRS azonban nagyon jól képes megkülönböztetni őket, a hibaarány 1 a 40 000-hez. Ez a kettős szűrő mechanizmusnak köszönhető: egy szűrő a túlságosan nagy amonosavakat szűri, míg egy másik a túlságosan kicsiket.32

Chaperonok
Amikor a riboszóma létrehozza a peptid láncot a fehérje még nincs kész. A sejt megfelelő működéséhez, beleértve az eddig tárgyalt enzimeket is, a fehérjéknek jól kell összetekeredniük, hogy kialakuljon a helyes háromdimenziós szerkezetük. A DNS szekvenciában kódolt végleges fehérjeszerkezetet a chaperonin nevű sejtgépezetek alakítják ki. Ezek olyan hordó alakú fehérjék, melyek más fehérjék összetekeredését segítik.33 Chaperonok nélkül például egy nagyon fontos fehérje hibás összetekeredése halálos priont eredményezhet. Ez okozza a Creutzfeldt-Jakob-szindróma nevű végzetes agykárosodást és a tehenek szivacsos agyvelőgyulladását (BSE) is, amit kergemarha-kórként is ismerünk.
A chaperonok is fékezik az evolúciós „folyamatokat”, mert gátolják a hibás fehérjék kialakulását, és így a változékonyság mértékét csökkentik. Egy új fehérje kialakulásánál szükségszerűen a szerkezete is megváltozik, és nagy az esélye annak, hogy összevissza hajlik vagy rosszul tekeredik. Egy ilyen fehérjének el kellene kerülnie a chaperonin szűrőket, amiknek épp az a feladata, hogy az ilyen hibásan tekeredő fehérjéktől megszabadítsa a sejtet. A chaperoninok minden élő szervezetben jelen vannak. Hogyan alakulhattak ki magasabb rendű szervezetek, ha nem voltak chaperoninok korábban? És hogyan jöhetett létre az első chaperonin, ha nem nem létezett egy ’elő-chaperonin’?34
A világ legkisebb motorja: az ATP-szintáz
Egy másik létfontosságú enzim az élőlényekben az ATP-szintáz, mely energiát biztosít az ATP-n (adenozin trifoszfát) keresztül. Az energia létfontosságú az élet számára, és minden élőlény ATP-t használ energiaforrásként.35 Minden élőlény, még a baktériumok és archeák is rendelkeznek ATP szintáz motorokkal.36 Ez teszi az ATP szintázt a leggyakoribb fehérjévé a Földön.
Valójában az emberi test minden nap a saját tömegével egyező mennyiségű ATP-t termel e motorok trillióival. Az ATP nagyon gyorsan felhasználódik a biokémiai reakciókban, beleértve a DNS- és fehérje szintézist, izomösszehúzódást, tápanyagok szállítását és az idegműködést. Egy élőlény ATP nélkül olyan, mint egy autó benzin nélkül. Bizonyos mérgek (pl. cián) az ATP termelés gátlásán keresztül hatnak. A szintáz az ATP-t a két komponens (ADP és foszfát) összekötésével hozza létre, energia felhasználásával. Az ATP-t ezután leválasztja, és felkészül az új ADP és foszfát fogadására. Ez a motor körülbelül 10 000 rpm-el forog és minden fordulatnál három ATP molekulát termel.
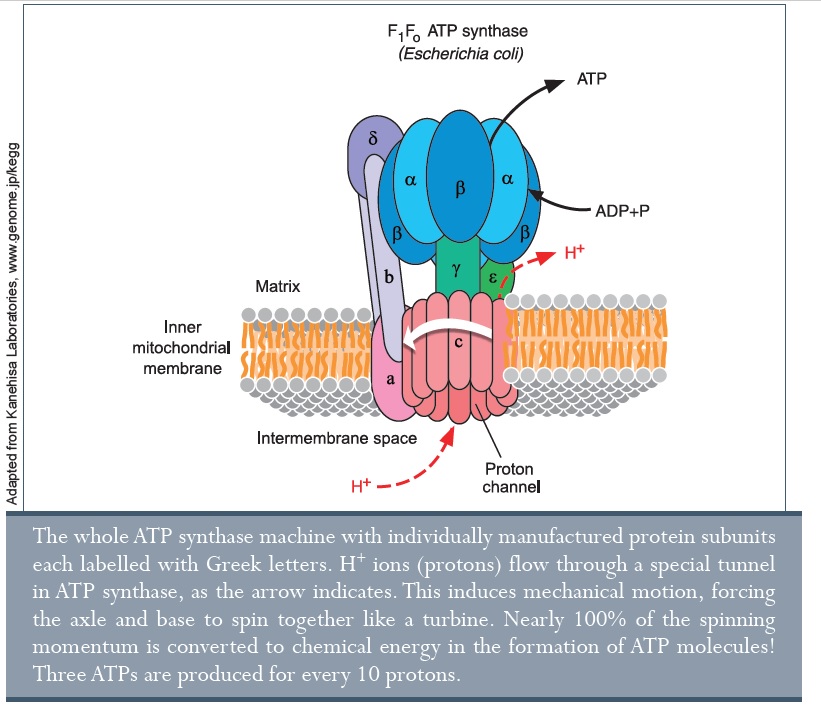
Valójában két motorból áll. A felső fele (F1-ATPáz) három részből áll, melyek mindegyike ATP-t termel. Az alsó felét (FO)37 a töltés-különbség hozza mozgásba, és pozitív töltésekkel (proton áramlás) működik, ellentétben az általunk épített elektromotorokkal, melyeket elektron-áramlás hajt.38 Időközben még egy részletre derült fény a motorok kapcsolódásával, ill. egymásra épülésével kapcsolatban.39 Jelenlegi kutatások azt mutatják, hogy ez a leghatékonyabb motor a világon – pontosabban, a fizikai törvények keretein belül a lehető leghatékonyabb. A kutatók következtetése szerint: „Eredményeink alapján az F1-ATPáz egy 100%-os energia-hatékonyságú mechanokémiai gépezet”.40 Vagyis egy kisebb csoda, hogy még a szigorúan világi Nature folyóirat is a „teremtés igazi motorjainak” nevezi őket.41
Enzimek
Az ATP-szintáz és az aminoacil-tRNS-szintetáz csak kettő a számos fehérje közül, amiket enzimeknek hívunk. Ezek a biológiai katalizátorok felgyorsítják a létfontosságú kémiai folyamatokat, anélkül, hogy maguk felhasználódnának. Az élethez rendkívül hatékony enzimek kellenek, különben a túlélés lehetetlen lenne, hiszen sok, az élethez elengedhetetlenül szükséges reakció túl lassan menne végbe, és nem is létezhetne az élet.42
Szuper katalizátorok
A különösen hatékony enzimek egyik jó példáját ismertette Richard Wolfenden 1998-ban.43 „A DNS és RNS felépülése nukleotidokból egy ’abszolút esszenciális’ reakció, mely 78 millió évig tartana vizes oldatban,44 de ez 1018-szorosára45 felgyorsul egy enzim révén.46 Ennek az enzimnek a működéséhez elengedhetetlen egy különleges, jól meghatározott szerkezet.47
2003-ban Wolfenden talált egy foszfatázt, amelyik a foszfát kötések hidrolizálását (hasítását) katalizálja. Ez az enzim a reakciót 1021-szeresére gyorsítja, ami még ezerszer gyorsabb az előző enzimnél! Ez az enzim lehetővé tesz a másodperc ezredrésze alatt végbemenő jelátviteli és szabályozási reakciókat az élő sejtekben. Enzim nélkül ez az esszenciális folyamat trillió éveket venne igénybe, ami százszor hosszabb idő, mint amit az evolucionisták az univerzum korának adnak meg (úgy 15 milliárd év)!48
Azonban ezek az enzimek és minden más folyamat, amiket az előbb felsoroltunk, mind jelen kellett legyenek az első szaporodó sejtben, hogy az túléljen és továbbadja a DNS-ét, és a DNS kell, hogy kódolja mindazokat a fehérjéket, amik ezekhez a folyamatokhoz szükségesek!
Következmények
Wolfenden szerint:
Katalizátorok nélkül nem létezne semmilyen élet, sem mikróbák, sem emberek. Nagyon nehéz elképzelni, hogy a természetes szelekció hogyan hozott létre egy olyan fehérjét, ami primitív katalizátorként működött volna egy ilyen rendkívül lassú reakció esetében.49
De ez egy érdekes helyzet. Ahogy korábban már rámutattunk, a természetes szelekció addig nem képes működni, amíg nincs élet, de az élet sem képes létezni enzimek nélkül, amelyek felgyorsítják az élethez elengedhetetlen reakciókat. Az élet létrejötte már az elején bukásra van ítélve a kémia alaptörvényei alapján!
Lehetséges egyszerűbb élet?
Ha ez még nem lenne elég, nem is annyira a kémia, mint inkább az információ a fő probléma az élet eredetével kapcsolatban. Még a legegyszerűbb elképzelhető élőlény működéséhez is rengeteg információ szükséges. A Mycoplasma genitalium baktérium a legkisebb genommal rendelkező ismert élőlény (a vírusokat nem számoljuk ide, mert a szaporodásuk nagyban függ a náluknál összetettebb sejtektől).50 A Mycoplasma genomja 482 gént tartalmaz, amit 580,000 bázispár kódol.51 Természetesen ezeknek a géneknek a működéséhez sejtmembrán és már létező transzlációs és replikációs rendszerek szükségesek. A Mycoplasma genitaliumnak azonban nincs sejtfala és csak parazitaként képes egy sokkal komplexebb élőlényben fennmaradni (pl. az ember légző- és urogenitális rendszerének sejtjeiben), ami biztosítja számára azokat a tápanyagokat, amiket saját maga nem tud előállítani. Valójában ez az élőlény annyira elvesztette genetikai információját, hogy a gazdaszervezettől függ a működése.52
A mycoplasmák nagyon egyszerű baktériumok (habár még így is bonyolultak). Létrejöhet ennél egyszerűbb élet? Mintegy tíz évvel ezelőtt Eugene Koonin kutató érdeklődött a mesterséges biológiai rendszerek iránt, és próbálta kiszámítani azt a minimumot, ami egy élő sejthez szükséges. Munkáját a mycoplazmákra alapozta, és azt próbálta megjósolni, hogy még mennyi gén híján tudnának működőképesek maradni. Csapata 256 génig jutott.53
Azonban kételkedtek abban, hogy egy ilyen hipotetikus élőlény sokáig életképes maradna, mert nem lenne képes a keletkező DNS-hibákat javítani, nem tudná finom-hangolni a meglévő gének működését, és nem lenne képes összetett vegyületek emésztésére sem, hanem komoly szerves táplálék-utánpótlásra szorulna.
Nem meglepő, hogy a további kutatások a szükséges gének számát sokkal többre becsülték. Az új hipotetikus minimális genom 387 fehérjét és 43 RNS-t kódoló gént tartalmaz.54
Egy 2009-es New Scientist cikkben ez szerepelt:
Nincs kétség afelől, hogy a közös ős DNS-ből, RNS-ből és fehérjéből épült fel, és egy univerzális genetikai kóddal rendelkezett, riboszómákat (fehérje szintetizáló egységek), ATP-t és proton hajtású ATP-szintetizáló enzimeket tartalmazott. A DNS-olvasás részletes mechanizmusa és a gének fehérjévé alakítása is már mind működőképes volt. Röviden, minden élőlény közös őse nagyon hasonlított egy mai modern sejthez.55
2011-ben evolúcióbiológusok értekeztek erről a hipotetikus közös ősről (angolul a LUCA rövidítéssel jelölték) és arra jöttek rá, hogy nem lehetett annyira egyszerű élőlény. Rendelkeznie kellett egy ’univerzális szervecskével’, ami pirofoszfatáz formájában energiát tárolt; egy korábbi dogma úgy tartotta, hogy a baktériumokból hiányzik ez az organellum.56 Ez állt a jelentésben:
A kutatók beszámolója szerint az új eredmények alapján LUCA egy meglehetősen bonyolult szervezet lehetett, összetett felépítése hasonlított a mai sejtekéhez.57
Az „élet eredete” szimulációk megközelítőleg sem ezt a minimumot veszik alapul.
Ki tud alakulni ilyen komplexitás véletlenszerűen?
A természetes szelekció csak az önfenntartó, szaporodó rendszerek esetén működik. Ezért nem lehet hivatkozási alap a minimális komplexitás kialakulásához. Az evolucionistáknak csak a véletlen marad. Ez nagyon egyszerű valószínűségszámítással kiszámolható.
Az információelmélettel foglalkozó Hubert Yockey kiszámolta, hogyha van egy medence, melyben biológiailag aktív aminosavak vannak (ami sokkal kedvezőbb kiindulási pont, mint a feltételezett „ősleves”), akkor az evolucionisták által feltételezett egymilliárd év alatt a teljes, létrejöhető információmennyiség nem több, mint egy 49 aminosav hosszúságú polipeptid.58
Ez körülbelül 1/8-a (információ tartalmát tekintve is) egy átlagos fehérje hosszának, de még a hipotetikus legegyszerűbb sejtnek is legalább 387 fehérjére van szüksége (DNS-ben kódolva!), és mindez csak egy speciális, változatlan, tápanyagokban és biomolekulákban gazdag környezetben jöhet létre. És Yockey nagyvonalúan számos olyan kémiai akadályt nem vett figyelembe, amit a későbbiekben még megnézünk, amik tovább nehezítik ezeket a folyamatokat.
Alternatív megoldásként ki lehet számolni a fenti fehérjékre vonatkozó DNS-szekvencia véletlen kialakulását. Természetesen van valamennyi mozgástér néhány fehérjénél, de nem az enzimek aktív centrumánál. Még az evolucionista szerzők is hallgatólagosan elismerik, hogy bizonyos szekvenciák esszenciálisak. Úgy nevezik őket, hogy „konzervált” szakaszok (azaz a szekvenciák annyira esszenciálisak (alapvető fontosságúak, nélkülözhetetlenek), hogy a természetes szelekció megőrizte őket az evolúciós folyamatban). Vannak konzervált fehérjék is, mint például a hisztonok, melyek a DNS kromoszómákba történő becsomagolásáért felelősek, vagy az ubiquitin, ami a baktériumokon kívül minden élőlényben megtalálható, és a nemkívánt, lebontásra kerülő fehérjéket jelöli59, vagy a calmodulin, univerzális kalcium-kötő fehérje, melynek majdnem mind a 140-150 aminosava „konzervált”.
A következő számítás nagyon nagylelkű lesz az evolucionisták számára. Azt feltételezzük, hogy enzimenként csak 10 aminosav helye konzervált, és létezik olyan mechanizmus, amivel az aminosavak oldatban is hosszú láncokat tudnak kialakítani (ez túlzottan nagyvonalú, mert oldatban a víz folyamatosan hidrolizálja a peptidkötéseket):
• 20 aminosav
• 387 fehérje a legegyszerűbb lehetséges élethez
• átlagosan 10 konzervatív aminosav
∴ ennek a valószínűsége: 20-3870 = 10–3870.log20 = 10-5035
Ez azt jelenti, hogy az egyest több mint 5000 nulla követi. És ez valószínűtlenebb, mintha valaki elsőre helyesen kitalál egy 5000 karakterből álló PIN kódot!60 Mégis, enélkül a teljesen véletlen összekapcsolódás nélkül nincs élet. Vagyis nem arról van szó, hogy lassan felépül a még nem élő vegyületekből az élő, hiszen ez az élet lehető legegyszerűbb formája azok szerint, akik azt hiszik, hogy az élet kialakulhatott egy vegyületekkel teli levesben.
Sok evolucionista azt mondja, hogy ha van elég idő, akkor bármi megtörténhet. Tényleg az idő a kulcsa mindennek? Nem. Nézzük csak:
• 1080 atom van az univerzumban
• 1012 atomi kölcsönhatás történik másodpercenként
• 1018 másodperc telt el az univerzum keletkezése, a feltételezett ősrobbanás óta
∴ így csak 10110 interakció lehetséges.
Ez egy hatalmas szám, de összehasonlítva a szükséges próbálkozások számával, azaz hogy a megfelelő, helyes nukleotid sorrend ki tudjon alakulni, ami képes a legegyszerűbb élethez szükséges fehérjék információját kódolni, abszurd módon kicsi. De még ilyen feltételek mellett is 104925 az esélye, hogy bekövetkezzen. Ezek a számok felfoghatatlanul hatalmasak. Ezek jól mutatják az élettelen vegyületekből kialakuló élet statisztikai lehetetlenségét (túl a „valószínűtlenségen”).
A híres kozmológus, Sir Fred Hoyle (1915-2001), elvetette ateizmusát mikor számításba vette ezt az abszurd módon kicsi valószínűséget:
Az élet élettelen anyagból való kialakulásának valószínűsége egy az olyan nagy számhoz, ami után 40.000 nulla áll … Ez pedig bőven elegendő, hogy Darwint és az egész evolúcióelméletet eltemesse. Nem lehet olyan ősleves sem ezen a bolygón, sem máshol, ahol véletlen folyamatokból élet alakulhat ki, az élet csakis tudatos intelligencia terméke lehet.61
Önreplikáló molekulák?62
Hogy Popper dilemmájára (lásd korábban) és az élet minimális komplexitására választ találjanak, néhány evolucionista olyan elmélettel állt elő, hogy léteznie kell egy olyan molekulának, ami képes mind katalitikus feladatokra, mind önmaga reprodukálására is. Két lehetséges válasz van, a nukleinsavak (RNS) és a fehérjék.
Azonban az evolucionistáknak be kell ismerniük, hogy az RNS-ek gyenge katalizátorok, a fehérjék pedig gyengén replikálódnak. Nincs olyan RNS enzim, melynek hatékonysága hasonló lenne az élethez szükséges enzimek hatékonyságához, amilyeneket Dr. Wolfenden is vizsgált, és amelyek nélkül az élet elképzelhetetlen lenne. Nincs értelme alternatív életformákról sem elmélkedni, hisz a mi bolygónkon található életet akarjuk magyarázni, és az élet dacol a naturalisztikus elméletekkel. Az evolucionisták beismerik, hogy a ribozimok (laboratóriumban előállított RNS enzimek) nem hatékony enzimek; soha nem képesek elérni az élethez elengedhetetlen katalitikus hatékonyságot. Még Dawkins is elismeri ezt:
Darwin a „kicsi meleg tavacska” elméletében gondolkodott az élet eredetének kulcsáról és ezt a fehérjék spontán megjelenésében látta, bár ez Darwin egyik kevésbé ígéretes ötlete volt. … De van valami, amiben a fehérjék kiemelkedően rosszak, és ezt Darwin figyelmen kívül hagyta. Teljesen reménytelen a replikálódó képességük. Nem képesek önmagukat lemásolni. Ez azt jelenti, hogy az élet keletkezésének kulcslépése nem magyarázható a fehérjék spontán megjelenésével.63
Ezen kívül még számos kémiai akadály is nehezíti mind az RNS, mind a fehérje alapú keletkezési modelleket. Sőt az egyik elmélet támogatója a másik legkeményebb kritikusa. De valójában mindkét félnek igaza van, és az élet spontán keletkezése kémiailag lehetetlen!
Az „RNS világ”
A nagyon népszerű RNS-alapú élet elmélete egészen 1967-ig nyúlik vissza, amikor is Carl Woese észrevette, hogy az RNS nem csak a replikálódni képes, hanem katalizálni is tud bizonyos folyamatokat.64 Thomas Cech és Sidney Altman egymástól függetlenül igazolta, hogy bizonyos RNS szekvenciák katalitikus hatással rendelkeznek. Az „RNS katalitikus tulajdonságának felfedezéséért” Nobel díjat is kaptak 1989-ben.65
A ribozimok felfedezése sok evolúcionistát vezetett az RNS világ feltételezéséhez. Úgy gondolták, hogy az első élet, ami RNS-re épült, nem csak másolódni volt képes, hanem olyan funkciókkal is rendelkezett, mint az enzimek. Richard Dawkins is támogatta néhányszor ezt az elképzelést.66
Azonban vannak problémák a RNS világ hipotézisével:67
• Az RNS egy nagyon komplex molekula. Nagyon nehéz elképzelni, hogy magától ki tud alakulni az őslevesben.
• Az RNS sokkal kevésbé stabil molekula, mint a DNS, pedig már a DNS is nagyon instabil (lásd lentebb).
• Bár a RNS is építőkövekből épül fel (nukleotidok), azok önmagukban is komplex molekulák, amik nem tudnak kialakulni az őslevesben. Még a laboratóriumi szintézisük is meglehetősen bonyolult.
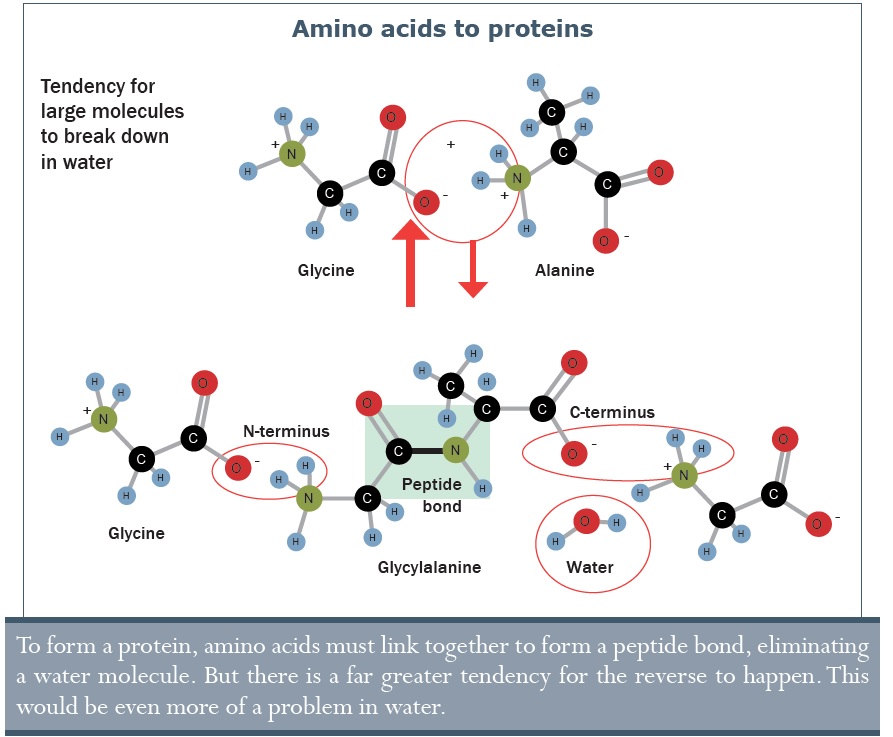
• Az elektromos kisülések, mint a Miller–Urey kísérletben, nem hoznak létre citozint, ami egyik alapköve a RNS/DNS-nek. A citozin, mégha létre is jönne, nagyon instabil ahhoz, hogy megfelelő mennyiségben halmozódjon fel, ugyanis a molekula felezési ideje 340 év 25 °C-on.68
• Még az RNS egyszerűbb építőkövei is instabilak a sejten kívül. A ribóz felezési ideje is mindössze 44 év 7,0 pH-n és 0 °C-on. És sokkal rosszabb magasabb hőmérsékleten (pl. 73 perc 100 °C-on).69 Nagyon gyorsan elbomlanak az RNS bázisok vízben 100 °C-on, ami egy nagy probléma a ’meleg tavacska’ vagy hőforrás elméletekkel.70 Az adenin és guanin felezési ideje egy év, az uracilé 12 év és a citoziné csak 19 nap.71
• Ahogy az aminosavaknál láttuk korábban, a nukleotidok sem képesek spontán polimerizálódni; aktiválódniuk kell. Továbbá, a polimerizáció számára optimális körülmények elősegítik ribozimok és a templátok lebomlását, amik viszont szükségesek a teljes folyamathoz.72 Még rosszabb, hogy a hidrolízis általános folyamata (a hosszú láncú biomolekulák bontása) is akadályozza a polimerizációt vizes közegben.73
• A kémiai reakciók által létrehozott molekulák királisak (tükörképei egymásnak). Bármilyen molekula, melyben a szénatomohoz négy különböző oldallánc kapcsolódik, lehet „bal kezes” és „jobb kezes” (lásd az ábrát), és bármilyen reakció az őslevesben csak 50:50 arányban hozza létre a molekulákat.74
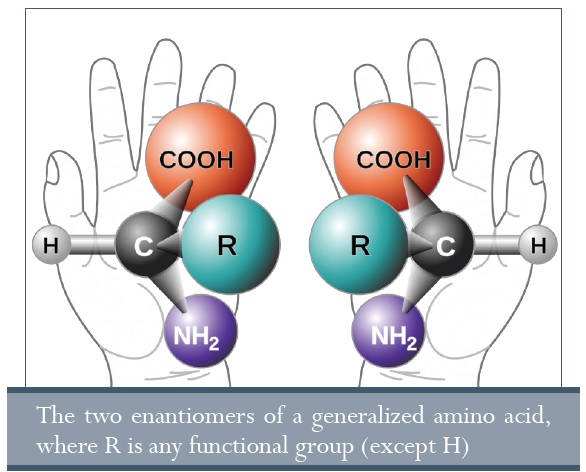 A nukleotid láncok spirálisan képesek összetekeredni (ez a stabilitás és replikáció miatt szükséges), de ehhez elengedhetetlen, hogy a molekulák „egy kezesek” (más szóval homokirálisak) legyenek. Mind az öt nukleotid bázis (A, C, G, T és U) jobb kezes, ahogy az összes cukor is a DNS gerincében. Érdekes, hogy az összes, élőlényekben található aminosav bal kezes. A homokiralitás miatt, ha csak egy molekula is rossz kiralitással kerül be az RNS szintézisekor, az megakadályozza annak másolását, még mesterséges körülmények között is.75
A nukleotid láncok spirálisan képesek összetekeredni (ez a stabilitás és replikáció miatt szükséges), de ehhez elengedhetetlen, hogy a molekulák „egy kezesek” (más szóval homokirálisak) legyenek. Mind az öt nukleotid bázis (A, C, G, T és U) jobb kezes, ahogy az összes cukor is a DNS gerincében. Érdekes, hogy az összes, élőlényekben található aminosav bal kezes. A homokiralitás miatt, ha csak egy molekula is rossz kiralitással kerül be az RNS szintézisekor, az megakadályozza annak másolását, még mesterséges körülmények között is.75
• Még ha ezek a polimerek létre is tudnának jönni, előzetes minta nélkül, önmaguk másolására is képessé kell válniuk. Ez a replikáció pedig, ha nem pontos, elvész az az információ, amit addig véletlenszerűen összegyűjtöttek. Azonban még ha 96,7%-os lenne is a pontosság, ami az eddig ismert legnagyobb pontosság,76 az sem lenne elég és hatalmas katasztrófát okozna. Az emberi DNS replikációjakor nagyon ritkák a hibák, ugyanis egy az egy milliárdhoz a másolási hibázások aránya, hála a jól tervezett hibajavító enzimeknek.
• Ha az első életformák önreplikáló RNS molekulák voltak, akkor azoknak minden olyan funkcióval kellett rendelkezniük, amik szükségesek egy élőlény működéséhez. De eddig az RNS esetében csak néhány kémiai reakció katalizálását figyelték meg.
• Ezek után hogyan lehetséges, hogy egy RNS alapú organizmus tovább fejlődik egy a DNS-t fehérje katalizátorokkal másoló élőlénnyé? Ez egy teljesen új dekódoló rendszert igényel77, és teljesen valószínűtlen a váltás az RNS alapú rendszerről egy DNS/fehérje alapú, információ tároláson alapuló rendszerre.
• A foszfát a nukleinsavak egyik alapvető alkotóeleme, a szabad foszfát ion (PO43–) tehát elengedhetetlen a reakciókhoz, de az RNS világ kutatói a tengerekben napjainkban mérhető koncentráció milliószorosát használják. A természetben a foszfát gyorsan kicsapódik kalcium (Ca2+) és magnézium (Mg2+) ionok jelenlétében, és így egy oldhatatlan szilárd anyag keletkezik, ami az összes ismert elméleti forgatókönyvet meghiúsítja.78
Nem csoda, hogy az „RNS világ” modellezésével kapcsolatos kutatások egyik vezetője, Gerald Joyce, ezt írta:
A legésszerűbb feltételezés az az, hogy az élet nem az RNS-el kezdődött. … Az átmenet az RNS világba, de ugyanígy az élet eredete általában, tele van rengeteg bizonytalansággal és hiányoznak hozzá a kísérleti adatok.79
Egy másik kémiai evolúcióval foglalkozó kutató, Robert Shapiro, miután kimutatta, hogy az RNS egyik építőköve valószínűtlen alkotóeleme az őslevesnek, ezt mondta:
A jelenleg rendelkezésre álló bizonyítékok nem támasztják alá azt az elképzelést, hogy RNS, vagy a jelenlegi RNS bázisokat használó replikátor jelen lett volna az élet keletkezésénél.68
Az élet eredetére vonatkozó erősen hiányos forgatókönyv fájdalmas „Achilles sarka” az evolúciós elméletnek. Bár nem csak ez az egyedüli probléma. Talán vannak még dolgok, amiket még mindig nem értünk. Amit azonban biztosan tudunk az az, hogy minden, amit a fizikáról, kémiáról tanultunk, teljesen valószínűtlenné teszi az élet keletkezését szervetlen vegyületekből.
„Fehérje volt először” elméletek
A kémiai evolúció régebbi elmélete szerint a fehérjék voltak az élet alapjai. És ez hatalmas reklámfelületet és lendületet kapott a híres Miller-Urey kísérletek kapcsán az 1950-es években.
Miller – Urey kísérletei
Stanley Miller (1930-2007), egyetemi tanítványa volt Harold Urey-nek (1893-1981), aki 1934-ben kémiai Nobel-díjat kapott a deutérium (nehéz hidrogén) felfedezéséért.80 Miller és Urey egy zárt üvegedény-rendszert töltöttek meg redukáló gázokkal: metánnal, ammóniával és hidrogénnel (és kizártak minden oxidáló gázt, mint amilyen az oxigén). A zárt rendszerben víz is volt, amit felforraltak és így a gőz cirkuláltatta a gázokat a 60 000 voltos szikrákon keresztül, amivel a villámokat szimulálták. A gázkeverék keresztül haladt egy vízhűtéses kondenzátoron és a reakciókban keletkezett vegyületek egy alsó ülepítőben gyűltek össze.
Egy hét után vöröses csapadékot találtak az ülepítőben. Az üledék oldhatatlan, toxikus és karcinogén volt, mely közönséges mellékterméke a szerves kémiai reakcióknak. De a csapadékban kis mennyiségben aminosavakat is találtak.81
Azonban ezek a legegyszerűbb aminosavak voltak, mint a glicin és az alanin.82 Az arányuk is minimális volt, mindössze 1,05% és 0,75%. Miller elismerte, hogy „a végtermék a felhasznált energiához képest nagyon csekély”.82 És nincs rá bizonyíték, hogy hosszabb idő alatt javulna a helyzet. Úgy tűnik, ez a szimuláció csupán csekély mennyiségű és egyszerű végterméket képes létrehozni.83
Ironikus megvizsgálni a feltételezéseket, az elvégzett kísérleteket, és az elkészített üvegberendezést, amivel ilyen minimális mennyiségű aminosavat tudtak csak előállítani, amiből nem keletkezhetett az élet; mégis fennen hirdetik, hogy „nem szükséges intelligencia az élet keletkezéséhez”!
Miller és Urey valójában azt fedezte fel, hogy véletlen kémiai reakciók random vegyületeket hoznak létre. Ugyanakkor ezek a vegyületek nagyon egyszerűek voltak. Egyszerű molekulák véletlen elegye pedig épp ellentétes azzal, ami az élet feltételeihez szükséges.
Kémia: az élet spontán keletkezésének halálos ellensége
A DNS, a sejt információ-tároló molekulája, egy nagyon instabil molekula. Egy új, a DNS stabilitásáról megjelent közlemény szerint a megőrződött csontokban, a DNS 22 000 év alatt esik szét teljesen 25°C-on, 131 000 év alatt 15°C-on, 882 000 év alatt 5°C-on, és 6,83 millió év alatt bomlik le teljesen -5°C-on.84 Egy másik cikk szerint:
„Általános meggyőződés, hogy a DNS „sziklaszilárd” – extrém stabil,” mondja Brandt Eichman, a biológia tudományok professzora Vanderbilt-ban, a projekt irányítója. „De a DNS valójában nagyon reaktív.”85
Ahogy a 2. fejezetben utaltunk rá, egy átlagos napon körülbelül egymillió bázispár sérül meg egy emberi sejtben. Ezeket a károsodásokat a sejten belüli normális kémiai aktivitások és a környezeti hatások kombinációja okozza, ilyen környezetből érkező hatás a sugárzások és a toxinok is, mint például a cigarettafüst, a grillezett ételek és az ipari hulladékok.86
A napi károsodások magas szintje miatt van az élőlényeknek DNS javító mechanizmusokra szüksége. A Chicago Egyetem biológusa, James Shapiro hangsúlyozza:
A baktériumtól az emberig, minden sejt rendelkezik egy igazán lenyűgöző javító rendszerrel, ami a véletlen és sztochasztikus mutációs forrásokat semlegesíti. A többszintű javítási mechanizmusok felismerik és eltávolítják azokat a hibákat a genomból, amelyek a DNS-replikáció során elkerülhetetlenül előfordulnak. … a sejtek megvédik magukat a véletlen genetikai változások azon típusaitól, amelyek a hagyományos elmélet szerint az evolúciós változatosság forrásai. Az ellenőrző és javító rendszereik működésének köszönhetően az élő sejtek nem passzív áldozatai a véletlen kémiai és fizikai erőknek. Jelentős energiát fordítanak a véletlen genetikai módosulások minimalizálására és képesek alacsony szinten tartani a környezet lokalizált mutagenitását a javító rendszereik aktivitásának szabályozása által.86
Természetesen, az elméleti ősleves nem tartalmazta ezeket a fantasztikus javító mechanizmusokat. Tehát ha a DNS képes is lenne spontán kialakulni, akkor sem maradna fent sokáig.
A RNS pedig sokkal bomlékonyabb, mint a DNS. Az RNS instabilitása az egyik fő ok, hogy sok kémiai evolucionista a fehérje alapú élet kialakulását támogatja. De ilyen hosszú idő alatt a fehérjék is éppen ennyire bomlékonyak és mindenképpen megsemmisülnek. Rájuk is vonatkozik a termodinamika második törvénye, és így az atomok véletlen mozgása és a háttérsugárzás miatt ők is lebomlanak végül. Például a víz képes lebontani a fehérjéket alkotóelemeire, aminosavakra (hidrolízis),87 és veszélyes keresztreakciók is történhetnek a fehérjék és más vegyületek88 között a feltételezett őslevesben.89 És a nukleotidokhoz hasonlóan, ahogy a korábbiakban volt erről szó, a fehérjéknél is bal és jobb kezes molekulák keveréke keletkezik (racemát) és nem homokirális vegyületek (csak jobb vagy csak balkezes formák), amik az élőlényekben található enzimekben megfigyelhetők.90
A jelenlegi vizsgálatok alapján a felső határ fagypontnál (0 °C) 2,7 millió év a kollagénnél és 110 millió év a csontfehérje (osteokalcin) esetében. Azonban már 10 °C-nál a felső határ már kevesebb, mint 180 000 év a kollagénnél és 7,5 millió év az osteokalcinnál. 20 °C-on a maximális élettartam még kevesebb: 15 000 és 580 000 év.91 Általában a hőmérséklet növekedése a reakciók sebességét növeli (biológiai reakciók esetében 10 °C-onként a sebesség megkétszereződik),92 és ez a probléma megoldhatatlan a meleg forrás elmélet esetében. A fehérjék instabilitása pedig erős érv a hosszú földtörténeti időszakok ellen: találtunk fehérjéket olyan dinoszaurusz csontokban, amelyek, ha valóban 65 millió évesek lennének, akkor nem lenne szabad megtalálnunk őket.93 További probléma az evolucionisták számára a szintén a dinoszaurusz csontokban talált DNS felfedezése is. A felfedezők kizárták a szennyeződés lehetőségét és a DNS annyira ép maradt, hogy kisméretű kettős hélixek is megfigyelhetőek voltak.94
Elfogadhatatlan vizsgálati beavatkozás
Az élet keletkezését igazoló szimulációk azt sejtetik, hogy csak időre és véletlenre van szükség. Valójában azonban a gyenge eredmények is csak a kísérletek beállítása miatt lehetségesek. Elfogadott, hogy ha A vegyületet megtalálják egy szikra kisüléses kísérletben, majd pedig B vegyületet megtalálják egy más típusú kísérletben (ami néha összeegyeztethetetlen az előző kísérlettel), akkor azt állítják, hogy „Lásd, az A és B képes létrejönni valós, ősföldi körülmények között.” Majd az iparilag mesterségesen előállított tiszta, homokirális és koncentrált formában lévő A és B vegyületből létrehozzák a bonyolultabb C vegyületet.95 Ezután pedig világgá kürtölik, hogy C vegyület létrejöhet az ősföldi körülmények között. De ez egyáltalán nem bizonyítja, hogy hígabb oldatban A és B képes reakcióba lépni egymással, vagy hogy nem lépnek reakcióba D, E vagy F vegyülettel, ami az első kísérletben, mint szennyeződés keletkezett. Röviden, az evolucionisták kísérleteiben az intelligens beavatkozás elfogadhatatlan szinten van.96
Az evolúciós propaganda többsége a következő feltételezett autó-keletkezési elmélethez hasonlít:
A tervezés egy tudománytalan magyarázat, ezért egy naturalista indoklást kell találnunk. Tehát a kísérletek azt mutatják, hogy az autó egyik fontos alkotóeleme (a vas) kinyerhető hevítéssel bizonyos ásványokból, mint például a hematitból, olyan hőfokon, amilyen néhol megtalálható a Földön. Ráadásul, a vas nagy nyomáson lemezzé formálható, amire bizonyos geológiai formációkban látunk példát. …
Ha az autómobil keletkezése túlságosan elnagyoltnak tűnik, akkor még vegyük figyelembe, hogy egy sejt sokkal nagyobb információ-tartalommal rendelkezik, mint egy autó, amiben nincs semmi, ami a belső homeosztázis fenntartásához szükséges és még szaporodni sem képes.
Élet versus ’piszkos’ kémia
A biomolekulák tulajdonságai nem az összetevőktől függnek, hanem az alkotórészek elrendeződésétől, vagyis az információtól. Ezek az elrendeződések nem az alkotóelemek fizikai és kémiai jellemzőiből származnak, hanem egy magasabb szintről. Hogy megerősítsük a fent említetteket, a szén, hidrogén, oxigén, nitrogén, foszfor és kén kémiai és fizikai tulajdonságaiból nem jósolható meg a makromolekula végső felépítése (hiszen végtelen a kombinációk lehetősége). Inkább úgy tűnik, hogy a makromolekulákat úgy tervezték meg, hogy a speciális funkcióikat az egyszerűbb alkotóelemek tulajdonságainak figyelembe vételével lássák el.
A makromolekulákat felülről lefelé tervezték, alulról felfelé nem alakulhattak ki. Az élőlényekben található gépezet a korábban bemutatott módon a megfelelő elemek rendezettsége alapján (információ) az alkotóelemekből önmaga másolatát képes elkészíteni, és ez így volt az előző generációban és azelőtt is. A gépezet nem az alkotóelemekből fejlődik ki, mert az olyan lenne, mintha zajból egy szimfónia jöhetne létre.
Minél inkább megismerjük a biokémiát, annál határozottabban tudjuk, hogy nem azonos az abiotikus kémiával. A törvények azonosak, de a sejten kívüli kémia mindig ’piszkos’ tömegreakciókból áll, míg a biokémia a tiszta, egyedi molekulák reakciója.97
A ’piszkos’ kifejezés a kémiai evolúcióval foglalkozó, Nobel-díjas Christian de Duve-tól származik. Tény, hogy az élőlényeken kívüli reakciókban sok vegyület vesz részt szabályozatlanul és sok szennyeződés van jelen.98 A teista evolúcióval foglalkozó paleontológus, Simon Conway Morris az élet eredetét modellező kísérletek termékeit „piszoknak”, „ragacsnak” és „trutyinak” nevezi,99 hasonlóan Graham Cairns-Smith, kémiai evolucionista, „erősen szennyezett trutyi” kifejezéséhez.100
Még a modern, ipari kémia, a legszigorúbb körülmények között sem képes 99,99%-nál nagyobb tisztaságot elérni. Ez főként a reakcióban egyszerre résztvevő molekulák hatalmas mennyisége miatt van. De a biokémia tisztább, mert az enzimek a reakciókban a molekulákat egyesével módosítják. A keletkező vegyületek ezután a megfelelő állapotban a megfelelő módon jutnak a következő enzimhez.
Végeredményben a sejtek kémiája pontos, szabályozott és kontrollált, csak így működőképes, és az élet másképp nem is létezne. A sejten kívüli kémia teljesen ellentétes azzal, ami az élethez és annak keletkezéséhez szükséges Az abiotikus kémia az élet antitézise. Hogy hozhatta volna létre ez az első élő sejtet?
Valószerűtlen csapdák
Minden energia, ami képes a biológiai vegyületeket létrehozni, az el is tudja őket pusztítani. Már említettük a víz hidrolizáló hatását, ami képes a peptidkötések bontására, és ez gyorsabban történik, mint ahogy a kötés létrejöttét segítené. Az energiaforrások is sokkal pusztítóbbak, mint amennyire konstruktívak. Például, a feltételezett ősi Földön az UV sugárzásnak két formája létezett. Az egyik a pusztító (ionizáló), rövidebb hullámú (UV) fény. A másik a nem káros, hosszabb hullámú UV. A hosszabb hullámú UV néhány esetben hasznos. Például a klorofill képes elnyelni és hasznosítani az energiáját.101 Azonban a káros UV-sugárzás gyakoribb, mint a konstruktív UV-sugárzás, ráadásul a hatása is nagyobb mértékű. A mértéke a felszínen kétszeres, pusztító hatása pedig körülbelül 104-105-szer erősebb, mint a konstruktív sugárzásé.102
A Miller-Urey kísérletekben ülepítőket terveztek bele a rendszerbe, hogy a keletkezett vegyületek elkülönítsék, azért hogy az elektromos kisülések (vagy az UV-sugárzás a későbbi kísérletekben) ne tegye tönkre őket. Ülepítő nélkül, még a keletkezett kis mennyiségű hasznos vegyület is elveszett volna. De ez nem valószerű modellezése az őslevesnek, mert nem volt olyan prebiotikus mechanizmus, ami az aminosavakat a káros UV-sugárzástól, az atmoszférától vagy a víztől elkülönítette volna. Jól tudjuk, hogy még egy gyengén felhős napon is képes az ember leégni, és még tíz méter mélyre is képes lejutni az UV-sugárzás a tiszta vízben.103 Tehát az ülepítők használata szintén egy olyan mesterséges beavatkozás, ami elfogadhatatlan ezekben a kísérletekben.
Kémiai evolúció: tény vagy vak hit?
A nem kreacionista, információ elmélettel foglalkozó Hubert Yockey, egy nagyon meglepő állítást tett 30 évvel ezelőtt:
Az élet eredetével kapcsolatos kutatások olyan szempontból egyediek, hogy a végső következtetést már határozottan elfogadták … . Ami még hátra van, hogy megtalálják azokat a forgatókönyveket, amik leírják a részletes mechanizmusokat és folyamatokat, ahogy ez lezajlott.104
Ezt fontos észben tartanunk, amikor népszerű evolúciós írásokat olvasunk, vagy amikor „elfogultsággal” vádolják a tervezettségben hívőket. Dr. Yockey így fejezte be cikkét:
Arra a következtetésre kell jutnunk, hogy a jelenleg kialakult ismerettel ellentétben az élet keletkezésének folyamatát a Földön, amit véletlennel és természeti okokkal magyaráznak, és tényekre, nem pedig hitre alapozva lehet elfogadni, még nem írták meg.105
Az Élet Eredete Alapítvány 1 millió dollárt ajánlott fel annak, aki kémiailag elfogadható naturalisztikus megoldást a genetikai kód és az élet keletkezésére.
Ezt olvashatjuk a honlapjukon:
Az „Élet-Eredete Díjat” (továbbiakban „a díj”) annak adjuk, aki javaslatot tesz egy nagy valószínűséggel működő mechanizmusra, ami képes megmagyarázni a genetikai kód spontán keletkezését a természetben, és elégséges magyarázat az élet keletkezésére is. A díj elnyeréséhez a magyarázatnak összhangban kell lennie a tapasztalati biokémiával, kinetikával és termodinamikai törvényekkel, továbbá az eredményt tudományosan elfogadott, szakmai folyóirat(ok)ban kell leközölni.106
De eddig nem voltak díjazottak, és minél többet tudunk meg az élet minimum követelményeiről, annál valószínűtlenebbnek tűnik, hogy az élet keletkezése elfogadhatóan magyarázható lehet materialista alapokon. A probléma talán soha nem került ennyire messze a megoldástól.107
Hol a bizonyíték?
Az előző levezetésből látszik, hogy az általános kémiai evolúció nem működik az őslevesben, de mégis kevesen veszik tudomásul, hogy nincs bizonyíték még az ősleves létezésére sem. A feltételezett ősleves az esszenciális nitrogén tartalmú aminosavak és nukleotidok forrása volt. Ha valaha létezett, akkor a geológusoknak találnia kellene olyan nagyon ősi kőzeteket, melyekben magas nitrogén tartalmú lerakódások vannak. Mégis alig találunk valamit, a legkorábbi szerves anyagnak nevezett kőzetben is alig van 0,015% nitrogén. Két geokémikus is kifejtette ezt:
Ha valaha volt ősleves, akkor azt várnánk, hogy a bolygónkon vannak olyan hatalmas üledékrétegek, melyekben különböző nitrogéntartalmú vegyületeket vagy azok maradványát találnánk, mint például savakat, purinokat, pirimidineket; vagy a metamorfózisos kőzetekben magas nitrogéntartalmat kellene mérnünk. De a tények azt mutatják, hogy ilyenek nincsenek a Földön.108
Élet az űrből?
A kémiai evolúció kudarcára reagálva egyes kutatók azzal érvelnek, hogy az élet nem a Földön jött létre, hanem az űrből származik. Ezt pánspermiának nevezik, a görög πάν (pán, minden) és σπέρμα (sperma, mag) szavakból alkotva, azaz az élet magja mindenhol jelen van az univerzumban.
A pánspermia elmélet klasszikus ága úgy tartja, hogy az élet magja természetes folyamatok, pl. üstökösök, által érkezett a Földre. Ez azonban nem nyújt megoldást, csak máshova helyezi a problémát. A kémiával és információval kapcsolatos nehézségek továbbra is megmaradnak. Amellett, hogy nagyon valószínűtlen, hogy bármi is túléli a millió évnyi utazást az űrben, hidegben, légkör nélküli és sterilizáló hatású kozmikus sugárzásnak kitéve. A legújabb kutatások azt mutatják, hogy a mikróbák nem képesek túlélni azt az extrém magas hőmérsékletet, ami az aszteroidákon a földi légkörbe lépéskor keletkezik – emiatt a hő miatt fénylenek a ’hulló csillagok’.109
Az újabb elméletet irányított pánspermiának nevezik, ami azt állítja, hogy földönkívüliek hozták az életet a Földre. Az elmélet támogatói között van a DNS kettős spirál egyik felfedezője Francis Crick (1916-2004) és Leslie Orgel is.110 Crick-et leginkább a kémiai evolúció elmélete zavarta, ezért pártolta ezt az új elképzelést:
Egy becsületes ember, minden jelenleg elérhető tudással felszerelkezve, arra az álláspontra kellene jusson, hogy az élet megjelenésének pillanata felér egy csodával, mert sok megfelelő körülményre volt szükség az élet elindulásához.
Minden alkalommal, amikor cikket írok az élet eredetéről, elhatározom, hogy soha többet nem fogok írni másikat, mert sokkal több benne a spekuláció, mint a tény.111
De újra meg kell jegyezni, hogy ez csak a probléma elodázása; el kell hinniük azt, hogy a hipotetikus űrlények is kémiai evolúció által jönnek létre az első bolygón.112 Érdekes, hogy azok, akik „tudománytalansággal” vádolják a teremtést, mert a Teremtő megfigyelhetetlen, elfogadják az irányított pánspermiát „tudományként”, a megfigyelhetetlen űrlények ad hoc elméletei ellenére. Meg kell említeni, hogy a pánspermia egy „hézagok evolúciója” elmélet, és az irányított pánspermia bizonyos tervezettséget is feltételez.
Összefoglalás
Az élet keletkezése az élettelen vegyületekből a vak hiten alapszik és nem a tudományon, és ez már Darwin ideje óta igaz. A legtöbb kémiai evolúciós elméletben a fő probléma az, hogy a teoretikusok a vegyületek együttesét tekintik életnek és nem az információfeldolgozó gépezet kialakulását. Soha nem teszik fel azt a kérdést, hogy „hogyan szerezte meg a molekuláris gépezet a saját programját?”
A természetes szelekció nem képes megmagyarázni az első élet kialakulását. Gépekre van szükség az információ feldolgozásához. De ez az információ tartalmazza a gépek építési útmutatóját is. Vagyis, máris itt a „tyúk vagy tojás” probléma. Ezek a gépek energiát használnak, amit az ATP-szintetáz motor állít elő. De a motort nem lehet megépíteni a DNS-ben kódolt utasítások és olvasó gépezetek nélkül, és ezeknek a molekuláknak is szüksége van ATP-re. Ez így már egy pete-lárva-szöcske probléma.
Más enzimek pedig nélkülözhetetlenek a létfontosságú reakciókhoz, melyek nélkülük csak évmilliók alatt mennének végbe. Az enzimek azonban önmagukban destruktívak is lehetnek, pl. egy izolált ATP szintáz elhasználja az ATP-t és nem termeli.
A fehérjék nem reprodukálódnak, és maguktól lebomlanának a természetben. A hosszú nukleotidok és poliszacharidok szintén lebomlanak (valójában létre sem tudnak jönni) a hidrolízis miatt. Az RNS is egy sérülékeny vegyület, sokkal bomlékonyabb mint a DNS. Az RNS néhány alkotóeleme nem jött létre a kémiai evolúciót „szimuláló” kísérletekben. A többi hígul, kicsapódik és/vagy elbomlik. Továbbá ezek sem „egy-kezesek” kiralitás szempontjából, ami az élethez elengedhetetlen.
Végül a legkorábbi kőzetekben nem találunk bizonyítékot a korai kémiai őséletre vonatkozóan.
Lényegében a fizika és kémia törvényei és a valószínűségek nem támogatják az élet abiotikus keletkezésének elméletét. A nem bibliai kreacionisták támogatják az isten-a-résekben érvelést! Sőt, végső következtetésünk és állításunk az, hogy a többszáz éves tudományos kutatások azt támasztják alá, hogy egy intelligens „teremtő Isten”, aki az univerzumon kívül létezik, az egyetlen ésszerű magyarázat az élet keletkezésére.
Hova vezet ez?
 Ez a könyv a darwini evolúció fő hajtómotorja, a természetes szelekció vizsgálatával kezdődött, amit hiányosnak találtunk. Ezután a genetikával foglalkoztunk, amit Darwin még figyelmen kívül hagyott, még ha szükséges is az evolúció megértéséhez, és azt láttuk, hogy az elmélet itt is elbukott. Ez a két fejezet jó párost alkot, mert ezek a témák központi szerepet játszanak Darwin elméletében. A jelen fejezet a következő, Dr. Emil Silvestru ősmaradványokat bemutató fejezetével áll párban. Ez a kettő olyan téma, melyeknek Darwint kellene igazolniuk, és tényleg ezeket terheli leginkább az elmélet igazolásának súlya, de akkor még a tudomány nagyon gyerekcipőben járt. Láttuk, hogy Darwin arra törekedett, hogy elkerülje az élet eredetére vonatkozó vitákat és erre jó oka volt! Kiderült, hogy Charles Darwin kerülte a fosszilis ősmaradványokkal kapcsolatos részletek megvitatását is, mert tudta, hogy nem támogatják az elméletét, ahogy ez teljes összhangban van napjaink tudásával. Ha az evolúció igaz, akkor először is a kémiának és a statisztikának igazolnia kellene (de nem), és másodjára az ősmaradványoknak is igazolnia kellene ezt. Darwinnak igaza volt, és a jövő majd igazolni fogja, hogy elég-e a millió éveken keresztül tartó lassú változás? De ki fog derülni, hogy az ősmaradványok éppen egy másik Akhilleusz sarka az evolúciós elméletnek.
Ez a könyv a darwini evolúció fő hajtómotorja, a természetes szelekció vizsgálatával kezdődött, amit hiányosnak találtunk. Ezután a genetikával foglalkoztunk, amit Darwin még figyelmen kívül hagyott, még ha szükséges is az evolúció megértéséhez, és azt láttuk, hogy az elmélet itt is elbukott. Ez a két fejezet jó párost alkot, mert ezek a témák központi szerepet játszanak Darwin elméletében. A jelen fejezet a következő, Dr. Emil Silvestru ősmaradványokat bemutató fejezetével áll párban. Ez a kettő olyan téma, melyeknek Darwint kellene igazolniuk, és tényleg ezeket terheli leginkább az elmélet igazolásának súlya, de akkor még a tudomány nagyon gyerekcipőben járt. Láttuk, hogy Darwin arra törekedett, hogy elkerülje az élet eredetére vonatkozó vitákat és erre jó oka volt! Kiderült, hogy Charles Darwin kerülte a fosszilis ősmaradványokkal kapcsolatos részletek megvitatását is, mert tudta, hogy nem támogatják az elméletét, ahogy ez teljes összhangban van napjaink tudásával. Ha az evolúció igaz, akkor először is a kémiának és a statisztikának igazolnia kellene (de nem), és másodjára az ősmaradványoknak is igazolnia kellene ezt. Darwinnak igaza volt, és a jövő majd igazolni fogja, hogy elég-e a millió éveken keresztül tartó lassú változás? De ki fog derülni, hogy az ősmaradványok éppen egy másik Akhilleusz sarka az evolúciós elméletnek.
Hivatkozások:
1. Névtelen forrás, az általános ősélőlény is sokkal komplexebb, mint korábban gondoltuk, ScienceDaily, 2012 október 3.; sciencedaily.com.
2. Lásd: Lamont, A., Louis Pasteur (1822–1895), Creation 14(1):16–19, 1991; creation.com/louis-pasteur.
3. Darwin, C., A fajok eredete, 1. kiadás, John Murray, London, 1859.
4. Levél Joseph Hooker-nek, 1863, Darwin, F. (szerk.), Charles Darwin élete és levelezése, vol. 3, p. 18, John Murray, London, 1887; elérhető a darwin-online.org.uk honlapon.
5. Darwin, F. (szerk.), Charles Darwin élete és levelezése, Vol. II, D. Appleton & Co., New York, pp. 202-203, 1911.
6. Dawkins legnépszerűbb érvelésének cáfolatát lásd Sarfati, J., The Great Hoax on Earth? c. könyvében, Creation Book Publishers, Powder Springs, GA, USA, 2010, elérhető a creation.com honlapon.
7. Habermas, G. interjúja Flew, A-val, Zarándoklatom az ateizmustól a teizmusig, Philosophia Christi, Winter 2004; biola.edu.
8. Flew, A., és Varghese, R., There is a God, Harper Collins, New York, 2007. Lásd még Cosner tanulmányát, L., J. Creation 22(3):21–24, 2008; creation.com/flew.
9. Johnson, P.E., Response to Gould, Origins Research 15(1):10–11, 1993; www.arn.org. Lásd még Q’Leary, D. dokumentumait: A darwinista Theodosius Dobzhansky nem volt orthodox keresztyén! post-darwinist.blogspot.com, 2006. szeptember 8.
10. Dobzhansky, T.G., quoted in Schramm, G., Discussion of Synthesis of Nucleosides and Polynucleotides with Metaphoric Esters, in: Fox, S.W. (ed)., The Origins of Prebiological Systems and of Their Molecular Matrices, pp. 309–310, 1963, Academic Press, New York, 1965.
11. Slack, G., What neo-creationists get right, The Scientist, 20 June 2008; the-scientist.com.
12. Kerkut, G.A., Implications of Evolution, Pergamon, Oxford, p. 157, 1960.
13. John Burdon Sanderson Haldane FRS (1892–1964) a legismertebb matemetikai populáció genetikus, de foglalkozott enzimekkel is Cambridge-ben, megalkotója a Briggs–Haldane enzime kinetikai törvénynek 1925-ben. Ő is mint határozott ateista és kommunista, támogatta a naturalista élet keletkezését lásd a Rationalist Annual folyóiratban, p.148, 1928.
14. Dickerson, R.E., Chemical evolution and the origin of life, Scientific American 239(3):62–102, 1978. szeptember
15. Pleasant, L.G. and Ponnamperuma, C., Chemical evolution and the origin of life, Origins of Life and Evolution of Biospheres 10(1):69–85, 1980.
16. Haeckel, E., The History of Creation, angolra fordította Prof. E. Ray Lankester, M.A., F.R.S, 3rd ed., Vol. 1, p. 184, Kegan Paul, Trench & Co., London, 1883. Lásd még Grigg, R., Ernst Haeckel: Evangelist for evolution and apostle of deceit, Creation 18(2):33–36, 1996; creation.com/Haeckel.
17. Bradbury, S., The Microscope Past and Present, Pergamon Press, 1968; van Niekerk, E., Countering Revisionism, J. Creation 27(1): 78–84, 2013; creation.com/haeckel-fraud-proven-part-2.
18. Dawkins, R., A vak órásmester, W.W. Norton, New York, p. 115, 1986.
19. Dawkins, R., A legnagyobb mutatvány, Free Press, New York, p. 405, 2009.
20. Polanyi, M., Life’s irreducible structure, Science 160:1308, 1968.
21. Orgel, L., The Origins of Life, John Wiley, New York, p. 189, 1973.
22. Davies, P., Life force, New Scientist 163(2204):27–30, 1999.
23. Williams, A., Life’s irreducible structure, J. Creation 21(1):109–115, 2007; creation.com/autopoiesis.
24. Popper, K.R., Scientific Reduction and the Essential Incompleteness of all Science, in Ayala, F. and Dobzhansky, T. (eds.), Studies in the Philosophy of Biology, University of California Press, Berkeley, CA, USA, p. 270, 1974.
25. Dawkins, The Greatest Show on Earth, p. 420.
26. Revyakin, A. et al., Abortive initiation and productive initiation by RNA Polymerase involve DNA scrunching, Science 314(5802):1139–1143, 2006; Kapanidis, A.N. et al., Initial transcription by RNA polymerase proceeds through a DNA-scrunching mechanism, Science 314(5802):1144–1147, 2006; lásd még az áttörés a DNS transzkripcióban a nanotechnológiai eszközökkel; physorg.com, 2006. november 16.
27. Roberts, J.W., RNA Polymerase, a scrunching machine, Science 314(5802):1139–1143, 2006.
28. Garrett, R., Mechanics of the ribosome, Nature 400(6747):811–812, 1999.
29. Truman, R. and Borger, P., Genetic code optimisation: Part 1, J.Creation 21(2):90–100, 2007; creation.com/gencode
30. Ibid.
31. Sarfati, J., Decoding and editing designs: double-sieve enzymes, J. Creation 13(1):5–7, 1999; creation.com/doublesieve; Karlson, P., (tr. Doering, C.H.), Introduction to Modern Biochemistry, 4th ed., Academic Press, London and New York, pp. 113, 145–146, 1975.
32. Nureki, O. et al., Enzyme structure with two catalytic sites for double-sieve selection of substrate, Science 280(5363):578–582, 1998.
33. Sarfati, J., DNA: marvellous message or mostly mess? Creation 25(2):26–31, 2003 március; Lásd még a ’DNS transzláció’ animációt a creation.com/message honlapon.
34. Aw, S.E., The Origin of Life: A critique of current scientific models, J. Creation 10(3):300–314, 1996; creation.com/origin-of-life-critique.
35. Bergman, J., ATP: The perfect energy currency for the cell, Creation Res. Soc. Q. 36(1):2–10, 1999; creationresearch.org.
36. Hiroyuki Noji et al., Direct observation of the rotation of F -ATPase, Nature 386(6622):299–302, 1997.
37. Jegyzet: az alsó indexben szereplő O jel, nem a nulla számot jelöli, hanem az ’oligomicin kötő rész’ rövidítését jelenti. Az oligomicin antibiotikum specifikusan blokkolja a protoncsatornát a bakteriális ATP szintáz FO részében, ami halálos.
38. Sarfati, J., Design in living organisms (motors), J. Creation 12(1):3–5, 1998; creation.com/motor. Lásd még Thomas, B., ATP synthase, Creation 31(4):21–23, 2009; creation.com/atp-synthase. Több részlet található még Sarfati, J., By Design c. könyvben, 10. fejezet, “Motors”, elérhető a creation.com honlapon.
39. Davies, K.M. et al., Macromolecular organization of ATP synthase and complex I in whole mitochondria, Proc. Nat. Acad. Sci. USA 108(34):14121–14126, 2011.
40. Toyabea, S. et al., Thermodynamic efficiency and mechanochemical coupling of F1 -ATPase, Proc. Nat. Acad. Sci. USA 108(44):17951–17956, 2011.
41. Block, S.M., Real engines of creation, Nature 386(6622):217–219, 1997 (perspective on Hiroyuki Noji et al., Direct observation of the rotation of F1-ATPase).
42. A katalizátorok nem változtatják meg az egyensúlyi állapotot, de annak elérését meggyorsítják. Alacsonyabb aktiválási energiával működnek, ami azt jelenti, hogy csökken az energiája az átmeneti állapotnak vagy a köztes reakcióknak. Lásd még az ábrákat és magyarázatot: Wieland, C. and Sarfati, J., Dino proteins and blood vessels: are they a big deal? 9 May 2009; creation.com/dino-proteins.
43. Miller, B.G. et al., Anatomy of a proficient enzyme, Proc. Nat. Acad. Sci. USA 97(5):2011–2016, 2000.
44. Ez azt jelenti, hogy 78 millió év kell a reakció felének végbemeneteléhez. A kémiában a reakciók fél életidejét használják a reakciók sebességének meghatározásához.
45. Idézet Lang L.H.-tól, Enzimek nélkül az ismert lassú biológiai reakciókhoz 1 trillió év kell: tanulmány, UNC School of Medicine 262:30, 2003; unc.edu. Lásd még: Wolfenden, R. and Snider, M.J., The depth of chemical time and the power of enzymes as catalysts, Acc. Chem. Res. 34:938–994, 2001.
46. Ez az enzim az ortodin-5’-monofoszfát dekarboxiláz, ami az uridin-5’-foszfát de novo szintéziséért felelős, ez esszenciális vegyülete a DNS és RNS bioszintézisének, ami az orotidin-5’-monofoszfát (OMP) dekarboxilációjával jön létre.
47. Több információ elérhető itt: Sarfati, J., World record enzymes, J. Creation 19(2):13–14, 2005; creation.com/enzymes; és itt: Sarfati, J, By Design, ch. 11, a creation.com honlapon keresztül.
48. Lad, C., Williams, N.H., and Wolfenden, R., The rate of hydrolysis of phosphomonoester dianions and the exceptional catalytic proficiencies of protein and inositol phosphatases, Proc. Nat. Acad. Sci. USA 100(10):5607–5610, 2003.
49. Lang, enzim katalízis nélkül a leglassúbb ismert biológiai reakciókhoz 1 trillió év szükséges.
50. Még a vírusok is rendelkeznek egy nano-motorral a DNS becsomagolásához Sarfati, J. Virus has powerful mini-motor to pack up its DNA, J. Creation 22(1):15–16, 2008; creation.com/virusmotor; Fuller, D.N. et al., Single phage T4 DNA packaging motors exhibit large force generation, high velocity, and dynamic variability, Proc. Nat. Acad. Sci. USA 104(43):16868–16873, 2007.
51. Fraser, C.M. et al., The minimal gene complement of Mycoplasma genitalium, Science 270(5235):397–403, 1995; perspective by Goffeau, A., Life with 482 genes, Science 270(5235):445–446. Más közlemények eltérő számokat közölnek, de a nagyságrend hasonló.
52. Wood, T.C., Genome decay in the Mycoplasmas, Impact 340, 2001; icr.org.
53. Wells, W., Taking life to bits, New Scientist 155(2095):30–33, 1997.
54. Glass, J.I. et al., Essential genes of a minimal bacterium, Proc. Nat. Acad. Sci. USA 103(2):425–430, 2006.
55. Lane, N., Was our oldest ancestor a proton-powered rock? New Scientist 204 (2730):38–42,2009.
56. Seufferheld, M. et al., Evolution of vacuolar proton pyrophosphatase domains and volutin granules: clues into the early evolutionary origin of the acidocalcisomes, Biology Direct 6:50, 2011.
57. Illinois Egyetemen az Urbana-Champaign-on, Névtelen, Az utolsó általános és közönséges ős sokkal bonyolultabb.
58. Yockey, H.P., A Calculation of the probability of spontaneous biogenesis by information theory, J. Theor. Biol. 67:377–398, 1977.
59. Truman, R., The ubiquitin protein: chance or design? J. Creation 19(3):116–127, 2005; creation.com/ubiquitin. Aaron Ciechanover, Avram Hershko és Irwin Rose nyerte a kémiai Nobel díjat 2004-ben „az ubiquitin közvetített fehérje lebontás felfedezéséért”; nobelprize.org.
60. Valójában a p érték nagyon alacsony, és d = 1/p, a számítás a következő: ahhoz hogy hogy 95% eséllyel sikerüljön egyszer a kísérlet, ahhoz 3 x d próbálkozás szükséges. Ebben a példában a p 10-5000, így d = 105000, tehát nekünk 3×105000 próbálkozás kell, hogy 95%-os eséllyel a szükséges enzimek létrejöjjenek. Dr. Jim Davidson személyes közlése (Észak-Karolina), 2012.
61. Idézve Major, E.L., Big enough to bury Darwin, Guardian (UK) oktatási anyagából, 2001 augusztus 23.; creation.com/hoyle-origin-of-life. Lásd még: Demme, G. and Sarfati, J., Big-bang critic dies, J. Creation 15(3):6–7, 2001; creation.com/hoyle.
62. Sarfati, J., Self-replicating enzymes? alapján J. Creation 11(1):4–6, 1997; creation.com/replicating.
63. Dawkins, The Greatest Show on Earth, pp. 419–420.
64. Woese, C., The Genetic Code, Harper and Row, New York, 1967.
65. Nyomtatásban: The 1989 Nobel Prize in Chemistry, 1989. október 12.; nobelprize.org.
66. Dawkins, A vak órásmester, 421.o.
67. Lásd még: Mills, G.C. and Kenyon, D.H., The RNA world: a critique, Origins and Design 17(1): 9–16, 1996; www.arn.org.
68. Shapiro, R., Prebiotic cytosine synthesis, Proc. Nat. Acad. Sci. USA 96(8):4396–4401, 1999.
69. Larralde, R., Robertson, M.P. and Miller, S. L., Rates of decomposition of ribose and other sugars: implications for chemical evolution, Proc. Nat. Acad. Sci. USA 92:8158–8160, 1995.
70. Sarfati, J., Hydrothermal origin of life?, J. Creation 13(2):5–6, 1999; creation.com/hydrothermal.
71. Levy, M. and Miller, S. L., The stability of the RNA bases, Proc. Nat. Acad. Sci. USA 95(14):7933–7938, 1998.
72. Johnston W.K. et al., RNA-catalyzed RNA polymerization, Science 292(5520):131925, 2001.
73. Sarfati, J., Origin of life: the polymerization problem, J. Creation 12(3):281–284, 1998; creation.com/polymer.
74. Sarfati, J., Origin of life: the chirality problem, J. Creation 12(3):263–266, 1998; creation.com/chirality.
75. Joyce, G.F. et al., Chiral selection in poly(C)-directed synthesis of oligo(G), Nature 310:602–604, 1984.
76. Johnston elismeri, hogy az általa létrehozott RNS-katalizált RNS polimeráz ribozim pontossága „még mindig elmarad a virális polymerázoktól melyek ≥0.996 pontossággal dolgoznak, de még ez is sokkal rosszabb, mint annak a polimeráznak a pontossága, ami a DNS másolását végzi.”
77. Further chemical problems are found in Cairns-Smith, A.G., Genetic Takeover: And the Mineral Origins of Life, Cambridge University Press, 1982; Lásd a honlap kivonatát: creation.com/rna.
78. Schirber, M., A salt-free primordial soup? Astrobiology Magazine, 2012. január 19.; astrobio.net.
79. Joyce, G.F., RNA evolution and the origins of life, Nature 338:217–224, 1989.
80. Harold C. Urey—Biography, nobelprize.org.
81. Miller, S. L., A production of amino acids under possible primitive earth conditions, Science 117:528–529, p. 528, 1953; Miller, S. L., Production of some organic compounds under possible primitive earth conditions, J. Amer. Chem. Soc. 77:2351–2361, 1955.
82. Yockey egy Nature (415(6874):833, 2002) cikkben azt állítja, hogy nem Stanley Miller volt az első. Már korábban Walter Löb (1913), Oskar Baudisch (1913), Edward Bailey (1922 és Harold Urey (1928, 1929) végzett hasonló kísérleteket. Yockey állítása szerint Miller csak korszerű elválasztási és vizsgálati technikákkal bővítette ki a korábbi kísérleteket, mint például kétdimenziós papírkromatográfiával. Ennek a technikának a jelentőségét éppen a szerves kémia professzorom javasolta. Egy válasz cikkben (Nature 416(6880):475, 2002) Jeffrey Bada és Antonia Lazcano védi Miller kémiai evolúciós eredményeit, de Löb nem tanúsított érdeklődést az eredmények iránt.
83. Bergman, J., Why the Miller–Urey research argues against abiogenesis, J. Creation 18(2):74–84, 2002; creation.com/urey.
84. Allentoft, M.E. et al., The half-life of DNA in bone: measuring decay kinetics in 158 dated fossils, Proc. Royal Society B 279(1748):4724–4733, 2012.
85. Newly discovered DNA repair mechanism, Science News; sciencedaily.com, 2010. október 5.
86. Shapiro, J.A., A third way, Boston Review, p. 2, 1997. február/március; Sarfati, J., New DNA repair enzyme discovered, creation.com/dna-repair, 2010. január 13.
87. Sarfati, Origin of life: the chirality problem.
88. Pl. az amino csoport (-NH2) az aminosavakban reakcióba lép a cukorban lévő karbonil csoporttal (O=C<), és ekkor egy víz molekula (H2O) szabadul fel és egy imin (HN=C<) csoport jön létre. Az imin nem hasznos csoport az élet számára, ezért Bergman szerint a Miller-Urey kísérlet nem magyarázza az abiogenezist.
89. Ha az ősleves ilyen nitrogén tartalmú molekulákat termelt, akkor ennek miért nincs nyoma a „legrégebbi” kőzetekben? Cf. Brooks, J. and Shaw, G., Origins and Development of Living Systems, Academic Press, London, UK and New York, 1973.
90. Sarfati, Origin of life: the chirality problem.
91. Nielsen-Marsh, C., Biomolecules in fossil remains: Multidisciplinary approach to endurance, The Biochemist, pp. 12–14, June 2002. Lásd még Doyle, S., The real ‘Jurassic Park’, Creation 30(3):12–15, 2008; creation.com/realjurassic és Thomas, B., Original animal protein in fossils, Creation 35(1):14–16, 2013; creation.com/ancient-protein.
92. Ezt nevezzük az Arrhenius-egyenletnek k = A exp(-Ea /RT), ahol a k az egyenlet konstansa, A pedig a hőmérséklet függő konstans (nevezik frekvencia faktornak is), exp az exponenciális függvényt jelöli, Ea az aktiválási energia, R az egyetemes gázállandó, és a T az abszolút hőmérséklet.
93. Sarfati, The Greatest Hoax on Earth?, pp. 204–208.
94. Schweitzer, M.H. et al., Molecular analyses of dinosaur osteocytes support the presence of endogenous molecules, Bone 52(1):414–423, 2013; lásd még Sarfati, J., DNA and bone cells found in dinosaur bone, J. Creation 27(1):10–12, 2013; creation.com/dino-dna.
95. Az evolucionista Cairns-Smith hasonló eredményekre jutott a tipikus „élet eredete” szimulációs kísérletek eredményeivel kapcsolatban (Genetic Takeover)
96. Thaxton, C.B., Bradley, W.L., and Olsen, R.L., The Mystery of Life’s Origin, ch. 6, Philosophical Library Inc., New York, 1984.
97. Williams, A., Life’s irreducible structure—Part 1: autopoiesis, J. Creation 21(2):116–122, 2007; creation.com/autopoiesis.
98. De Duve, C., Singularities: Landmarks on the Pathways of Life, Cambridge University Press, Cambridge, 2005. Lásd Williams A. összefoglalóját: Great minds on the origin of life, J. Creation 21(1):38–42, 2007; creation.com/singularities.
99. Conway Morris, S., Life’s Solution: Inevitable humans in a lonely universe, Cambridge University Press, Chs 3–4, 2003. Lásd még ReMine, W. összefoglalóját: Evidence for Message Theory, J. Creation 20(2):29–35, 2006; creation.com/lifes-solution.
100. Cairns-Smith, A.G., Genetic Takeover and the Mineral Origins of Life, Cambridge University Press, New York, 1982.
101. A fotoszintézis egy másik olyan egyszerűsíthetetlen komplex rendszer ami négy foton energiáját tárolja és vele víz molekulákat képes bontani. Lásd Sarfati, J. cikkét: Green power (photosynthesis): God’s solar power plants amaze chemists, J. Creation 19(1):14–15, 2005; creation.com/greenpower.
102. Hulett, H.R., Limitations on prebiological synthesis, J. Theor. Biol. 24:56–72, 1969.
103. UV-B sugárzás 65 méter mélyre lehatol a tiszta antarktiszi vizekben: W.C Gieskes és G.W. Karay Transmission of ultraviolet light in the Weddell Sea: Report on the first measurements made in Antarctic, Biomass Newsl. 12:12–14, 1990.
104. Yockey, A Calculation of the probability of spontaneous biogenesis, p. 379.
105. Yockey, A Calculation of the probability of spontaneous biogenesis, p. 396.
106. Az Élet Eredete Díj; www.us.net/life.
107. Smith, Calvin, Who wants to be a millionaire, creation.com/lifeprize, 2007. aug. 15.
108. Brooks, J., and Shaw, G., Origins and Development of Living Systems, Academic Press, London and New York, 1973.
109. Meteorite experiment deals blow to bugs from space theory, Physorg.com, 25 September 2008; Sarfati, J., Panspermia theory burned to a crisp: bacteria couldn’t survive on meteorite, 10 October 2008; creation.com/panspermia.
110. Crick, F. and Orgel, L.E., Directed Panspermia, Icarus 19:341–346, 1973.
111. Crick, Life Itself, Its Origin and Nature, pp. 88, 153, Simon and Schuster, 1981.
112. Lásd még Bates, G., Designed by aliens? Creation 25(4):54–55, 2003; creation.com/aliens.
Ábrák
Könyvben 89. oldalon lévő – a honlapon 1. sz. ábra
Aminosav kötőhely
tRNS Akceptor szár
D hurok TΨC hurok
extra hurok
Antikodon
kodon mRNS
A genetikai kód működése: Három specifikus nukleotid az mRNS-en (kodon) hozzákapcsolódik a nukleotid párjához a tRNS-en (antikodon). Mivel a tRNS-ek a különböző antikodonokkal különböző aminosavakat szállítanak, az mRNS nukleotid szekvenciája meghatározza az aminosavak sorrendjét a fehérje végtermékben. Az egyes tRNS-hez az aminoacil-tRNS-szintetáz köti az aminosavakat ATP felhasználásával (nincs ábrázolva). creation.com/gencode
Könyvben 90. oldalon lévő – a honlapon 2. sz. ábra
Tyr
Phe Aktiváló hely.
A nagyobb aminosavak
nem jutnak át
Val
Ala
Gly
Ile
Hibajavító hely.
Val
Ala
Gly
Ile tRNS
Kisebb aminosavak nem jutnak át Az izoleucin-tRNS szintetáz kettős szűrő mechanizmusa. Egy hidrolítikus módosítás csökkenti a rossz valin aminosav beépülésének hibalehetőségét 1 a 10-ről, 1 a 100-ról, 1 a 40 000-re.*
*Fersht, A.R., Sieves in sequence, Science 280 (5363):541 (válasz Nureki et al. Enzyme structure with two catalytic sites for double-sieve selection of substrate cikkére), 1998.
Könyvben 92. oldalon lévő – a honlapon 3. sz. ábra
A teljes ATP szintáz nanomotor egyedileg előállított fehérje alegységekből épül fel, melyeket görög betűk jelölnek. A H+ ionok (protonok) egy speciális csatornán haladnak keresztül, ahogy a nyíl mutatja. Ez indítja be a mechanikai mozgást, a tengely és az alap együttes, turbina-szerű pörgését. A mozgási energia közel 100%-a alakul át kémiai energiává, ATP molekulák formájában! 10 proton után 3 ATP molekula termelődik.
Könyvben 99. oldalon lévő – a honlapon 4. sz. ábra
A fehérjékben az aminosavak peptid kötésekkel kapcsolódnak össze, és a reakció során vízmolekulák keletkeznek. De a fordított folyamat sokkal gyakoribb. Ez az igazi probléma a vízzel.