EAH 4. Silvestru: Ősmaradványok
Részlet az Evolúció Akhilleusz sarkai (halálos pontjai) című könyvből

4. fejezet:
Ősmaradványok
112-153 oldal
A könyvet Robert Carter, Ph.d szerkesztette.
Kiadó: Creation Book Publishers. Második kiadás, 2014. október
Honlap: Creation.com
Fordította: Konkoly Dávid.
Lektorálta: Dr. Hecker Kristóf biológus.
Dr Don Batten Ph.D: Az evolúció Akhilleusz sarkai című könyvének fejezetenkénti fordítása.
Tartalomjegyzék:
1. fejezet: Természetes szelekció
6. fejezet: Radiometrikus kormeghatározás
8. fejezet: Etika és erkölcsiség
4. fejezet
Dr. Emil Silvestru
A geológia doktora,
Babes-Bolyai Egyetem, Románia
Dr. Silvestru tudományos képesítését a kommunista Romániában szerezte, ahol a barlangi geológia és a karszt-topográfia világszintű szakembere lett. Cikkei széles körben jelentek meg ezen a titokzatos szakterületen, és ő volt a világ első barlangkutató intézetének igazgatója Kolozsváron, Romániában. Emil később keresztény lett, és a föld koráról, ill. az evolúcióról alkotott korábbi nézeteit elkezdte felülbírálni. Természetes módon adódik tehát a választás, hogy ő kapja a darwinista evolúció negyedik Achilles sarkáról szóló fejezetet a kövületekről.
Lásd: creation.com/dr-emil-silvestru
Az ősmaradványok
Dr. Emil Silvestru, geológus
(Babes-Bolyai Egyetem, Románia)
Ha a napjainkban tapasztalható élet biológiai sokszínűsége a közel 4 milliárd éve zajló természetes szelekció eredménye, akkor az ősmaradványok nyújthatnák ehhez a végső bizonyítékot. Úgy tűnik, hogy egy efféle naturalista világnézet gyengeségeire nehezen találunk tényszerűbb példát, amely az evolúció valódi Achilles sarkává teszi a kövületeket.
Mi az a fosszilis lelet?

A „lelet” szó használata a „fosszilis lelet” kifejezésben azt sugallja, hogy a fosszíliák valamilyen formában megőrzik a történelmet, azonban ez jelentősen eltér azoktól a történelmi leletektől, amelyeket az ember őriz meg.
A Darwinisták számára a fosszilis leleteknek az a rendeltetésük, hogy bemutassák a fajok időben fokozatos evolúcióját. Ősidők óta tudott dolog, hogy egyes kőzetek bizonyos élőlények maradványait tartalmazzák, míg mások kövületekben szegények. A Brit-szigetek hatalmas területén található rétegek egész sorozatán lezajlott szisztematikus vizsgálat során – amelyet William „Strata”1 Smith végzett el a 18. század végén – alakult ki a következetes fosszilis sorozat koncepciója.
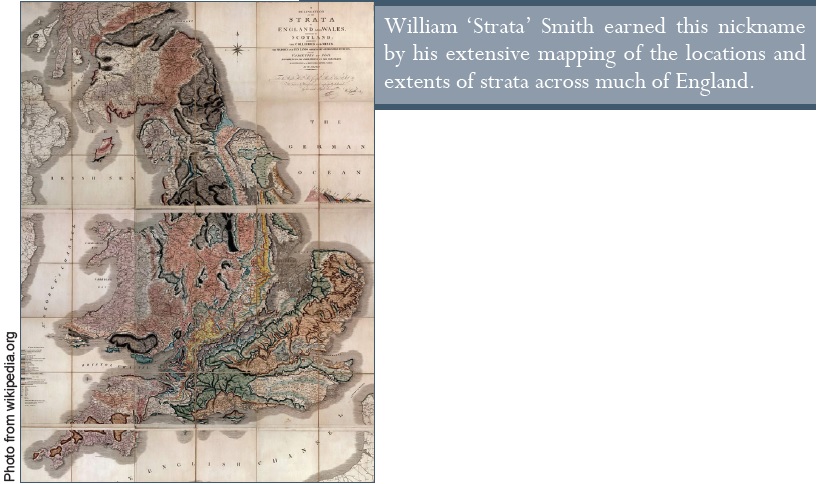
Képaláírás: William „Strata” Smith becenevét annak köszönheti, hogy részletesen feltérképezte Anglia szinte teljes területén a rétegek (lat. strata) helyét és kiterjedését.
Egy évszázadon belül a lelőhelyek egész világra kiterjedő hálózata jött létre, mint a napjainkban is működő Global Boundary Stratotypes Section and Point (GSSP). Ezek a pontok megkísérlik a kövületeket tartalmazó kőzeteket egyetlen földi rendszerbe illeszteni, ahol a kövületekre úgy tekintenek, mint amelyek komplexitásukban folyamatos és következetes növekedést mutatnak (az „egyszerűtől” az „összetettig”) a legidősebbtől a legfiatalabb üledékeken át. A tudományos világban a kőzet és a fosszilis leletek összhangba hozására való törekvés biostratigráfiaként ismert. Ennek a vizsgálata végeredményben a modern taxonómiának a múltba való extrapolálása. Habár ez elég ártalmatlannak tűnik, az alapvető evolúciós feltételezések alapján találták ki és fejlesztették tovább, hogy a későbbi taxonok a régebbiekből származnak. Így a vizsgálódás iránya kifejezetten evolúciós következtetéseket von maga után.
A taxonómia csupán a jelenlegi és a múltbeli organizmusok csoportosítására való törekvést jelent a legalapvetőbb közös tulajdonságaik alapján. Tudományként Carl Linné vezette ezt be, aki kreacionista volt. A filogenezis a taxonómia egy ága, amely olyan fához hasonló struktúrákat állít fel, amelyek arra hivatottak, hogy megjelenítsék az élőlénycsoportok közötti kapcsolatokat a taxonómiai adatok segítségével. Az eredmények bibliai és evolúciós kontextusban is értelmezhetőek, azonban a fa-ábrákat a vizuális megjelenésük okán gyakrabban használják evolúciós érvelésekhez. A paleontológia a tudománynak az az ága, amely a kövületeket tanulmányozza. Az adatok itt is bármelyik szempontból felhasználhatók, de mivel a paleontológusok többsége evolucionista, az ezen a területen megjelenő értelmezések evolúciós talapzaton állnak.

|
1. |
Domén |
Eukarióták |
5. |
Rend |
Főemlősök |
|
2. |
Ország |
Állatok |
6. |
Család |
Emberfélék |
|
3. |
Törzs |
Gerinchúrosok |
7. |
Nemzetség |
Hominini |
|
4. |
Osztály |
Emlősök |
8. |
Faj |
Homo sapiens |
1. táblázat: Az emberi faj besorolása a Linné-féle rendszertan szerint: Eukarióták: ezek a szervezetek maggal rendelkező sejtekből állnak;
Állatok: vagyis nem növények;
Gerinchúrosok: pálca alakú támasztó szervvel rendelkeznek (ebben az esetben gerinc);
Emlősök: melegvérű, szőrzettel rendelkező állatok, amelyek szoptatják a kicsinyeiket;
Főemlősök: majomszerű élőlények a magasabbrendűség hangsúlyával. Emberfélék: emberszerű élőlények;
Hominini: ember;
Homo sapiens: gondolkodó ember.
Mit várt Darwin a fosszilis leletektől?

Carl Linné a 18. században rakta le a modern taxonómia alapjait a kettős nevezéktannak egy olyan rendszerét használva, ahol minden fajnak egy kétszavas latin elnevezése van. Például az embert Homo sapiens-nek nevezik („ember” + „gondolkodó”). Linné óta a Földön élő, névvel ellátott fajok száma folyamatosan nő, amelynek egyelőre nem látjuk a végét. Bolygónk biológiai sokszínűsége továbbra is lenyűgöz minket. De min nyugszik ez a sokszínűség? Több millió állatfaj létezik, de csak 36 törzs! Sok faj van kevés számú törzsben, amely ugyanúgy igaz a növényekre, a gombákra, a baktériumokra és az archeákra. Más szavakkal, viszonylag kevés „alap formaterv” adja a földön lévő összes élőt. Ez tudott volt Darwin idejében, így látható, hogy ő a törzsek eredetét a múltba történő extrapolálásukkal képzelte el. Fajok sokasága tömöríthető kevés nagy csoportba, amelyek aztán különböző törzsekbe illeszthetők, amelyek egyetlen ősi formába (vagy formákba) tehetők (Darwin mind az egyedülit, mind a többest is említi).
Természetesen, ha a molekuláktól az emberig tartó evolúció lefedi bolygónk történetének nagy részét, és ha az üledékes kőzetek mindezen korszakok során egymásra rakódtak, akkor az üledékek az evolúció megfelelő lenyomatait kell(ene) hordozzák a fosszíliák számát és formáját tekintve, különösen az átmeneti fosszíliák esetében, amelyek összekötik az egymás után következő evolúciós fokozatokat. Pontosan ez az, amit Darwin várt a fosszilis leletektől, és csak így érthető meg csalódottsága, amikor be kellett vallania, hogy:
…ha a fajok más fajoktól észrevehetetlenül kis fokozatok során fejlődtek ki, miért nem látunk mindenhol számtalan átmeneti formát? Miért nem zűrzavaros a természet az egymástól tisztán megkülönböztethető fajok helyett, amint látjuk is?2
De jó darwinista szokás szerint elveti a problémát néhány bekezdéssel később:
…én itt csak azt állítom, hogy hitem szerint a válasz az, hogy a leletek nem annyira teljesek, mint ahogy általában feltételezzük; a leletek tökéletlensége jórészt abból adódik, hogy a szerves létezők nem a tengerek legmélyén tartózkodnak, valamint beágyazódott és konzerválódott maradványaik a későbbi korokra csak olyan üledékhalmokban képesek ellenállni a lebomlás hatalmas mértékének, amelyek kellőképpen tömörek és kiterjedtek; és ilyen üledék csak akkor halmozódhat fel, ha sok üledék rakódik le a tenger sekély fenekén, miközben lassan leülepszik.
Számos tévedés van ebben az állításban, ideértve abbéli hitét, hogy nem létezett élet a tengerek mélyén, amelyről nem sokkal ezután bizonyosodott be, hogy nem így van. Láthatjuk, hogy frusztrációja ma még nagyobb lenne, mivel magyarázata gyengének bizonyult.
Amire a bibliai kreacionisták számítanak
Mivel a kreacionista geológia középpontjában a Teremtés Könyvében található Özönvíz áll, az ebben hívő geológusok az alábbi várakozásokat támasztották a fosszilis leletekkel kapcsolatban:
1. Az ősmaradványok, amelyek javarészt az Özönvíz során elpusztult élőlények, hirtelen jelentek meg, általában előfutárok nélkül, egy adott ideig léteztek, majd utódok nélkül tűntek el, amennyiben ma nem találhatók élve.
2. Az összes főbb taxon (országok, törzsek, osztályok és rendek, esetleg egyes családok és nemzetségek, valamint elszigetelt, egy-taxonos fajok) közvetlenül az Özönvíz előtt jelen voltak, alaptípusként léteztek, amelyek aztán az Özönvíz után sok változatban (fajban) terjedtek szét.
3. A fosszíliák vertikális rendszerének vissza kellene tükröznie élőhelyüket és az árvíz dinamikáját (mind a vízmozgás, mind az élőlények formáját, méretét és viselkedésüket illetően). Ennek a rendszernek nem kell univerzálisnak lennie, sőt, a katasztrofális áradás és betemetődés eredményeként az egy helyen élő organizmusokat esetenként olyan élőlényekkel együtt kellene megtalálni a kövületben, amelyek teljesen más élőhelyről származnak.
4. A fosszíliák többségét a leletekben tengeri organizmusoknak kell(ene) kitennie.
5. Kevés vagy semennyi átmeneti forma nem létezhet, mivel az összes típus (nem faj) már meg volt teremtve, és jelen volt az Özönvíz kezdeténél. Lehetséges néhány nehezen besorolható forma (például a kacsacsőrű emlős) és olyan is, amelyet két kategóriába is lehet sorolni (ilyenek az „emlős-szerű hüllők”), de továbbra is jellemző a főbb átmeneti formák általános hiánya. Nem korlátozhatjuk Isten kreativitását, tehát ezt az állítást általánosságban értjük.
6. Kell, hogy legyenek olyan megkövült lábnyomok szárazföldi élőlényektől az Özönvíz korai szakaszából, amelyek vagy úszni tudtak vagy a lebegő növényzeten megmaradtak. Ezek olyan területeken érhettek földet, amelyeket időszakosan szárazra kerültek a dagályok, szökőárak, viharos hullámzások, Coriolis-erők (amelyeket a Föld forgása okoz) hatására vagy akár a kontinentális, ill. regionális tektonikus lemezek mozgásától is.
Ezután vizsgáljuk meg, hogy a fosszilis leletek mi módon igazolják ezeket a várakozásokat.
A fosszilis lelet, mint egy jelkép
Szinte úgy tűnik, hogy a kövületek mára egyfajta jelképpé váltak, egy olyan szentképpé, melyet lelkes követők tömegei részesítenek tiszteletben. Amikor az „aranyrudak”3 listáját olvassuk, amelyeket a fanerozoikus4 kőzetek geológiai korának meghatározására használtak, látjuk, hogy mennyi kövületre hivatkoznak úgy, hogy azok megfeleljenek az elvárásoknak, téve mindezt az ezeket cáfoló állítások sorozatának ellenére. Továbbá, mint minden szimbólumot, az ősmaradványokat is olyan szenvedélyesen védelmezik, hogy semmilyen paleontológiai tény ne kérdőjelezhesse meg nagyszerű és győzedelmes előrehaladását az időben. A gyakorlatban ez olyan szintre jutott el, hogy gyakran a paleontológusok szabják meg, hogy a kövületnek hol kell lennie ebben a folyamatban, és ha máshol találják meg, azonnal kimagyarázzák. Minden új és kihívást jelentő leletet beleerőltetnek, hogy passzoljon a róla kialakult képhez, míg azokat az új felfedezéseket, amelyekről úgy tűnik, hogy erősítik a paradigmát, diadalmasan népszerűsítik.
Ha megnézzük, hogy manapság a tankönyvekben és a médiában bemutatott fosszilis leletek hogyan fedik fel az élet történetét, gyakran könnyedén észrevehetünk bennük nagy rejtélyeket, amint azt a néhai Stephen Jay Gould világosan megfogalmazta:
(1) A soksejtű élet miért jelent meg olyan későn? (2) És miért nincs ezeknek az anatómiailag komplex élőlényeknek [Gould a Burgess-pala faunájára utal] közvetlen, egyszerűbb felépítésű előfutáraik a prekambriumi időkből5 származó fosszilis leletekben?
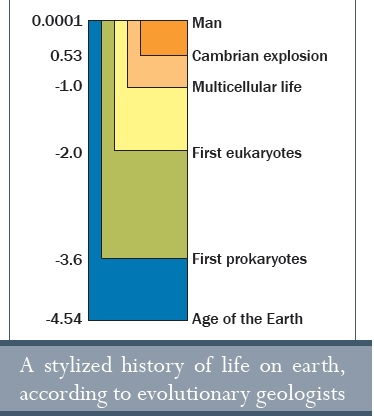
Időábra:
Ember
Kambriumi robbanás
Többsejtű élet
Első eukarióták
Első prokarióták
Föld kora
A földön lévő élet egy stilizált története az evolucionista geológusok szerint
Valóban, a többsejtű élet a föld feltételezett 4,54 milliárd éves evolúciós idejének csak egy töredékét reprezentálja. Mivel a legidősebb ismert létformáról úgy tartják, hogy mintegy 3,4 milliárd évvel6 ezelőtt keletkezett, a többsejtű élet az életnek csak egy részét mutatja be. Furcsa módon, ha valaki figyelembe veszi az evolúciós események időbeliségét, úgy tűnik, hogy minél komplexebbé vált az élet, annál gyorsabban evolválódott, de aztán ez az ütem drámaian lelassult, amint a főbb formák kifejlődtek.
A Darwini folyamatokkal kapcsolatosan szkeptikus evolucionisták csoportjáról készült ismertető írása közben Walter J. ReMine rámutatott:
Stephen Jay Gould megjegyzi, hogy a leletek sorrendje azt mutatja, hogy a legsokfélébb (legkülönbözőbb) biológiai struktúrák jelentek meg először! Ezeket követik a kevésbé eltérő felépítettségek. Ezeket pedig a még kevésbé eltérőek. Amíg végül az utolsó, fajok közötti biológiai sokszínűség legcsekélyebb jelét mutató formák mutatkoznak a folyamat legvégén.7
Azt várhatnánk, hogy az evolúciós korszakok alatt a fajok lassan elválnak egymástól és végső soron magasabb taxonómiai kategóriákat hoznak létre. Ehelyett a legnagyobb különbségeket felmutató kategóriák (az országok és a törzsek) jelennek meg először. Az egyértelmű tények kifejtése után (az evolúciós várakozás gyakran ellentéte annak, amit a kövületek mutatnak), ReMine így folytatja:
De ez egyre súlyosabbá válik. A genetika legújabb felfedezései egy újabb érdekes kihívást szolgáltatnak a problémához. Fejlődésbiológusok megfigyelték a géneknek egy olyan kis csoportját, amely hozzájárul a testfelépítés szervi kifejlődéséhez – és ez jelen van a többsejtűek világán át a különböző törzsekben és osztályokban. Az evolucionisták ezt „genetikus fejlődési eszköztárnak” nevezik. Az evolucionista gondolkodás szerint ennek a komplex eszköztárnak valamilyen közös őstől kell származni az össze törzs irányába. De ennek a közös ősnek a törzsek megjelenése előtt már léteznie kellett – más szavakkal a kambriumi robbanás előtt (amikor a fosszilis leletek sorában valamennyi törzs teljesen kifejlődve és hirtelen jelenik meg, evolúciós előfutárok nélkül). A közös ősnek (amelynek kiléte a mai napig ismeretlen) a prekambriumi időben kellett léteznie – a többsejtű élet keletkezése előtt. Röviden tehát a testfelépítés irányításáért felelős gének eredetének olyan időre kell visszanyúlnia, amikor még nem volt test. Az embriológiai fejlődést kontrolláló géneknek pedig olyan időre, amikor még nem voltak embriók.
Ez nagy nehézség azok számára, akik az élet naturalisztikus eredetében hisznek! Amint arra a 3. fejezetben már rámutattunk, a Végső Közös Ősnek (LUCA) kétségkívül nagyon komplexnek kellett lennie. De hol van a LUCA-ra vonatkozó bizonyíték? Nem található meg a kőzetekben, sőt, a genetika is csavar egyet ezen az evolúciós sztorin. Egy evolucionista, Michael Syvanen felfedezte, hogy nagyon sok eukarióta gén (minden, ami összetettebb, mint a baktériumok) megfelelője egyszerűen nem található meg a baktériumok génjei között.8 Ez nem lehetett igaz.
A félrevezető ábrákkal és szövegekkel mindezen tények homályosak maradnak azon tanulók számára, akikben az az összkép áll össze, hogy az élet evolúciójának folyamata egyenletes és kiegyensúlyozott ütemben zajlik. Az ezt bemutató ábra, mely az élet fájaként ismert, egy újabb szimbólum, amely egy manipulatív analógiát alkalmaz. Ami ezen a fán látható, az a törzs és az ágak. Azonban néhány evolucionista szerint nem ez a helyzet, és hogy a fa inkább olyan, mint egy kusza bokor. Erre az a magyarázat, hogy a genetika felfedezte, valójában nincs is fa. Ehelyett különféle organizmusok különböző géneken osztoznak, látszólag véletlenszerűen. Ez nem illik bele a darwini „életfa” koncepcióba; nem egyezik azzal az elképzeléssel, hogy minden faj egy közös ősön keresztül evolúciós összeköttetésben áll egymással; ezért az evolucionisták a horizontális génátvitelre (HGT) hivatkoznak. Ez az, amiről úgy hiszik, hogy ősi (bakteriális) fajok osztoztak egymással különböző DNS-szakaszokon és ez vezetett közvetlenül a fa gyökerének teljes összekuszálódásához.9
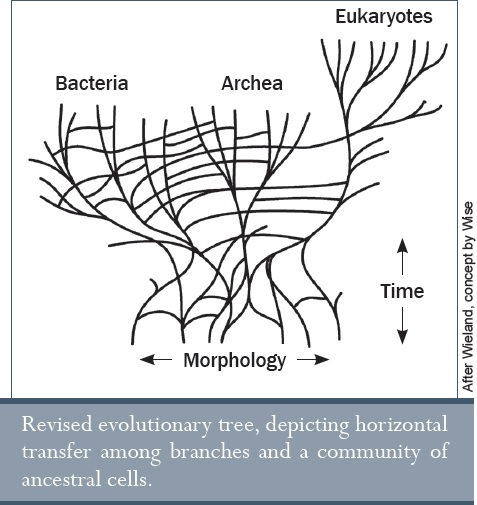
Ábra: Baktériumok – Archeák – Eukarióták – Morfológia – Idő (Wieland után, Wise koncepciója alapján)
Módosított evolúciós fa, amely az ágak közötti horizontális génátvitelt és az ősi sejtek közösségét ábrázolja.
Érdemes megjegyezni, hogy Darwin felhasználta az életfa analógiáját a „Fajok eredeté”-ben, bár ő inkább szőlőként vázolta fel (lásd 1. fejezet). Mivel a fosszilis leletek a múltbeli életnek az üledékes kőzetekben megőrződött archívumaként tekinthetők, így ennek evolúciós interpretációja szükségszerűen azoknak a felfedezéseknek a folyományaként alakul ki, amelyek az élő dolgok komplexitásának növekedését írják le a geológiai időtávon belül. Még pontosabban, annak a 36 állattörzsnek, amikkel ma a taxonómia foglalkozik, egészen az élet azon első formájáig visszakövethetőnek kell lennie, amelyről úgy hiszik, hogy élettelen összetevők természetes okai során jöttek létre. Azonban már nagyon korán hatalmas réseket fedeztek fel egy ilyen egyenes irányú leszármazási sor felállításakor, a kőzetekben talált fosszíliák pedig átírták az evolúciós eredetre vonatkozó elméletüket, gyakran az eredettörténet jelentős felülvizsgálatát okozva ezzel.
Néhány őskori lelet az evolúciós történet olyan alapos revízióját igényli, hogy erős késztetés irányul egy másodlagos hipotézis felállítására annak érdekében, hogy a paradigma megtartásával magyarázható legyen a helyzet. Ezeket a fosszíliákat gyakran a „származtatott” vagy az „átdolgozott” címkével látják el. Én inkább különálló fosszíliák-ként tekintek rájuk, értve ezalatt, hogy nem ott találták meg, ahol elvileg kellett volna, mert bizonyos ismeretlen (vagy feltételezett) geológiai folyamatok eltávolították őket eredeti (vagyis „set”) helyzetükből és átrakta őket jelenlegi (vagyis „offset”) helyükre.
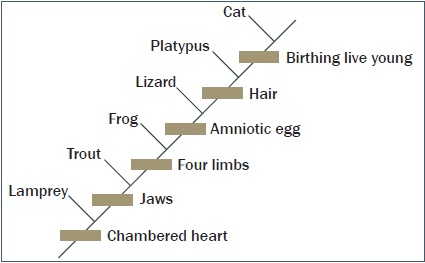 Lehetetlenné vált ilyen fát felállítani a rengeteg ág miatt, és azért is, mert a törzs egyes részei teljesen hiányoznak. Ehelyett kladogramokat használnak.10 Az evolucionisták a megosztott és a meg nem osztott jellegzetességek egymásba illesztett rekeszeit rajzolják fel, és ezekből fa-diagramokat ágaztatnak. Azonban ezek a sok paleontológiai tényből levont elképzelések titkos ábrázolásai! Eképpen egy kladogram tartalmazhat olyan fantázianevekkel ellátott csomópontokat, amelyek felfedezetlen élőlény-kategóriát jelölnek! Az itt bemutatott ábra az élet eredetének egy egyszerűsített elágazó fa-diagramja, amely számos forráson és gondosan kidolgozott kladogramon alapul.
Lehetetlenné vált ilyen fát felállítani a rengeteg ág miatt, és azért is, mert a törzs egyes részei teljesen hiányoznak. Ehelyett kladogramokat használnak.10 Az evolucionisták a megosztott és a meg nem osztott jellegzetességek egymásba illesztett rekeszeit rajzolják fel, és ezekből fa-diagramokat ágaztatnak. Azonban ezek a sok paleontológiai tényből levont elképzelések titkos ábrázolásai! Eképpen egy kladogram tartalmazhat olyan fantázianevekkel ellátott csomópontokat, amelyek felfedezetlen élőlény-kategóriát jelölnek! Az itt bemutatott ábra az élet eredetének egy egyszerűsített elágazó fa-diagramja, amely számos forráson és gondosan kidolgozott kladogramon alapul.
Ábra: cat: macska, platypus: kacsacsőrű emlős, lizard: gyík, frog: béka, trout: pisztráng, lamprey: orsóhal, chambered heart: üreges szív, jaws: állkapocs, four limbs: négy végtag, amniotic egg: tojásmagzat, hair: szőr, birthing live young: elevenszülés
Számos következtetést vonhatunk le a fosszilis leletekből. Először is, ami korántsem folytonos, a komplexitás időben való növekedése (ha lezajlott egyáltalán) szakaszos és hirtelen. Másodszor, az állatoknak sok olyan csoportja van, amelyeknél hosszú geológiai időszakok alatt sem látszik a fejlődés, ez a jelenség az evolúciós sztázis. A legmakacsabb példa a sztromatolitoké: ezek cianobaktérium kolóniák, amelyek nem változtak az állítólagos 3,4 milliárd év alatt, kivéve, hogy a legösszetettebb kolóniaformáik épp a legősibb múltban voltak jelen!11 Harmadszor, a legjelentősebb evolúciós lépések azokban az élőlényekben mentek végbe, amelyeket még nem fedeztek fel (evolúció a résekben!). Negyedszer, ha kövületeket találnak az evolúció logikai rendszerén kívül, a teljes evolúciós történet összedől. Richard Dawkins egyszer megjegyezte, amit már előtte J.B.S. Haldane is hangoztatott:
Azonban, ha csak egyetlen víziló vagy nyúl is létezett a Prekambriumban, ez teljesen romba döntené az evolúciót. Mindeddig semmi ilyesmit nem találtak.12
Vízilovakat és nyulakat lehet, hogy nem találtak a Prekambriumban, de tudja mit? Sok más olyan hasonló ősmaradványt találtak, amelyek evolúciós jelentőséggel bírnak. Ezek „romba döntötték az evolúciót”? Nem, mert az evolúció egy világnézet, amelyet arra használnak, hogy mindenféle bizonyítékot értelmezzenek vele, bármit is találjanak. Nem cáfolható egy fosszilis lelettel. A kövületet egyszerűen újraértelmezik, hogy úgy tűnjön, teljes mértékben bizonyítja az evolúciót.13
Most pedig elemezzünk néhány jól-dokumentált esetet, amelyet „eltérő kövületeknek” hívok, de mielőtt ezt tesszük, foglaljuk össze: a fosszilis leletek nyújtotta „összkép” egyfajta megszakítottságot és pangást mutat. Ezek a tények sokkal inkább megfelelnek egy kreacionista magyarázatnak.
„Eltérő kövületek”
A pakisztáni sóhegység
A Karakorum hegység lábánál, az Indus és a Jhelum folyók között, észak-kelet Pakisztánban elterülő sóhegység sok vitát generált a 19. század óta. A sóüledékek a Kehwra Homokkő (korábban Lila Homokkőként ismerték) alatt találhatók, amelyek Kambrium-korabeliek (állítólag 542-488 millió évesek).14 1927 elején a sóüledékekben növényi maradványok jelenléte keltette fel a geológusok figyelmét.15 Mivel a megkövesedett növényi részek újkori tölgyből származtak, a sóhegységet a Harmadkorra datálták.16

Kép: A sóhegység Pakisztán Punjab tartományában
1944-ben Sahni17 kövületek egész sorát említette, amelyeket a sórétegekben találtak: zárvatermő fa (virágos növény) foszlányait, nyitvatermő (nem virágzó növények, mint amilyen a fenyő) tracheidákat (folyadéktároló edények) és legalább egy jó, szárnyas, hatlábú, összetett szemmel rendelkező rovart. Találtak ősmaradványokat a só dolomit rétegeiben is. Sahni ezek korát az Eocénre tette (feltehetően 56-34 millió évvel ezelőtt, a dinoszauruszok kihalása után). Mégis a számos geológus által tett részletes helyszíni vizsgálatok azt mutatták, hogy a Kehwra Homokkő korát tekintve a Kambriumra tehető és az alatta fekvő sóhegység normális helyzetben van (vagyis nincs elcsúszva), ezért prekambriumi korúnak kell lennie.18 Sahni meglehetősen óvatos és szigorú vizsgálatokkal folytatta, miközben egy elszenesedett fát talált egy dolomit-tömbben. Be is mutatta az új fosszíliáknak (fa szövet) dolomitból való kinyerését egy geológiai konferencia résztvevői előtt Nagy-Britanniában.19 Azzal kapcsolatos erőfeszítése, hogy ezt a rendhagyó (offset) stratigráfiai helyzetet egyfajta réteggyűrődésnek tulajdonítsa, megbukott, mivel megkérdőjelezte volna a geológia alapelveit, ezért bevallotta:
Nemrégiben Mr. Gee egy alternatív magyarázatot kínált. Azt vetette fel, hogy a sólerakódások nyitvatermői, zárvatermői és rovarai a kambriumi és prekambriumi időszak magasan fejlett flórájának ill. faunájának a képviselői! Más szavakkal: azt feltételezik, hogy ezek az állatok és növények a sóhegység területén többszáz millió évvel korábban bukkantak fel, mint tették ezt bárhol máshol a világon. Alig hiheti bárki is, hogy egy ilyen gondolatot komolyan előterjesztene valaki ma a geológusok közül.20
Az 1950-es években tovább folytatódtak a sóhegység kövületeiről szóló beszámolók. Majd hirtelen csend lett, és bár az újabb vizsgálatok ezt az alakzatot az Eokambriumra teszik (kései Prekambrium),21 mindössze egyetlen olyan tanulmányt találtam, amely megemlíti a Khewra sóbarlangok fatöredékeinek felfedezését, de helytelen módon elveti és teljesen figyelmen kívül hagyja Sahni erős bizonyítékát az in situ helyzettel kapcsolatban.22 Jellemző, hogy egy ennél frissebb, 2009-ben publikált geológiai tanulmány, amely a fosszilis lelőhelyek védelmét ösztönzi, a sóhegységet prekambriumi korúnak írja le, de a kövületekről semmilyen említést nem tesz!23
A venezulai Roraima képződmény
1966-ban Stainforth bejelentette, hogy virágport fedezett fel a Roraima képződményben, amely egy olyan kvarc homokkő, amelyből a híres venezuelai tepuik – táblahegyek – állnak.24 A híres Arthur Conan Doyle képzeletbeli Elveszett világ-ának helyszíneként ismert és a világ legnagyobb vízeséséről (Angyal-vízesés) is híres lenyűgöző vidék ugyanakkor az evolucionisták számára rossz hírű Stainforth felfedezése miatt.

Kép: Angyal-vízesés, Venezuela
A képződmény „egybehangzóan” elfogadott, radiometrikusan meghatározott evolúciós kora 1,8 milliárd év, amely a Paleo-Proterozoikum egy olyan időszakának felel meg, amikor az evolúciós forgatókönyv szerint, zöld (fotoszintetizáló) növények még nem is léteztek! Kétségbeesett próbálkozásokat tettek ennek a problémának25 a kimagyarázására, mindhiába. Stainforth következtetése teljesen helyénvaló, miszerint „Erre az ellentmondásra nem tudunk megoldást nyújtani, így ez továbbra is komoly fejtörést okoz.”
Az Ediakara gerinces
A dél-ausztráliai Flinders Range-ben található Ediacara városa a többsejtű organizmusok „legkorábbi” (késő prekambriumi) gyűjtőhelye, amelyek jelentősen eltérnek az (evolúciós időskála szerint) újabbaktól. Habár az első ehhez a korhoz és életfához tartozó kövületet, az Aspidella terranovica-t Újfundlandon találták 1868-ban, mégis Ediacara-ban találták meg e furcsa élőlények egész sorát. Ma már a késő prekambriumi időszakot ediakarainak nevezik. A kövületek többségét olyan cső- és tollformájú gerinctelenek képviselik, amelyek láthatólag nem rendelkeznek modern megfelelőkkel.
Majd 2003-ban különböző hírforrások bejelentették a legkorábbi gerinces (gerinchúros) felfedezését ugyanabból a leletcsoportból.26 Úgy tekintették, mint „az élet fájának legalsó része”,27 az élőlénynek nem volt neve és minden további nélkül szinte egy pillanat alatt eltűnt! Miután ez némi felbolydulást keltett, a tudományos közösség annyira tanácstalan volt, hogy tudatosan nem említették ezeket a leleteket. Nem úgy, mint más helyzetekben, amikor megtaláltak egy kihívást jelentő „offset” fosszíliát, a „mainstream” tudományos média nem volt elárasztva leleplező cikkekkel. A kövületeket alapvetően azóta is figyelmen kívül hagyják, amely egy dolgot jelenthet: ez egy valós kihívás az élet eredetének evolúciós forgatókönyve számára. Ez azért van, mert az elmélet az első gerincesek evolúcióját a jóval távolabbi múltba helyezi, indokolatlanul rövid időt hagyva az ős-gerinchúrosok evolúciója számára. Ami még súlyosbítja a dolgot, hogy a fosszíliákra vonatkozóan nincs semmi, ami összekötné a gerinceseket a korábbi időszakokkal. Úgy tűnik, hogy a gerincesek minden észrevehető vagy egyáltalán elképzelhető ős nélkül léptek az élet színpadára!

Kép: A Dickinsonia costa nevű ediakarai őskövület
Edényes növények a Kambriumban
A paleobotanika általános nézete ma az, hogy az első szárazföldi edényes növények a Felső Ordovíciumban (450 millió évvel ezelőtt) fejlődtek ki, holott már régen ismertek azok a kelet-szibériai leletek, amelyek szerint ezek már a Kambriumban léteztek (540 millió évvel ezelőtt).28 Hasonló bizonyítékot találtak Svédországban és Észtországban.29 Mivel korábban már találtak pollent ős-prekambriumi sziklákban és erezettel rendelkező növényeket a késő Prekambriumban, a Kambriumhoz köthető edényes növényeknek nem kellene meglepetést okozniuk, hacsak ez nem azzal járna, hogy a növények evolúciója jóval korábban kezdődött, ill. hogy fosszíliákban közismerten szegény üledékes kőzetleletekkel kell foglalkozni. Ezek az őskövületek hirtelen véget vetnek a sajtó által szívesen és gyakran felkapott ősklíma-hipotézisnek, a Hólabda Földnek,30 ami a jéggel borított bolygót a Felső-Prekambriumra helyezi. Valamilyen módon ez idézte elő – amint rejtélyes véget ért közvetlenül a Kambrium előtt – az élőlények valódi robbanását a formák megdöbbentő sokszínűségével.

Ábra: Evolúciós időskála (millió év) Geológiai oszlop
Nem szokványos környezetben fosszilizálódott állatok
Az „offset” fosszíliáknak van egy másik kategóriája, amiről ritkán beszélnek: állatok maradványai, amelyek teljesen más környezetben éltek, mint amiben megkövesedtek.
1. Hadroszauruszokat (kacsacsőrű dinoszauruszok) találtak Montana „Medvemancs” palájának tengeri üledékeiben. Habár 1900-ban fedezték fel és 1902-ben publikálták, a paleontológusok döntő többségének figyelmét 1979-ig elkerülte (szárazföldi lények tengeri üledékben eltemetve; ez nem pont az, amit vártak!). Napjainkban ezeket a felfedezéseket továbbra is ritkán említik, holott egyre több ilyen lelet kerül elő, amely alapvetően kizárja a nyílt tengeren való hirtelen betemetődést.
2. Nodosaurus (páncélozott dinoszaurusz) fosszíliákat találtak fejjel lefelé tengeri üledékekben (beleértve a mészkövet) betemetve Nyugat-Kansasban.32 Az elsőre a híres O.C. Marsh bukkant rá 1870-ben, aki az Apatosaurus felfedezője volt.33
3. 2011-ben egy Ankylosaurus csontvázat fedeztek fel kátrányhomokban az albertai Fort McMurray mellett;34 egy újabb nagy meglepetés, mivel a homok tengeri üledék, amely már korábban számos tengeri hüllő (mosaszaurusz) kövülettel szolgált.
Ez csak néhány a rengeteg példa közül. Ezeknek a felfedezéseknek a gyakorisága és széles földrajzi kiterjedése inkább egy hatalmas vízi katasztrófára utal, mintsem a tetemek többszáz kilométeren át tartó véletlenszerű szállítására a nyílt tengeren.35 Ez különösen igaz, mivel még a nagyméretű tetemeket is, csontokkal együtt, sok tengeri élőlény hamar elfogyasztja, amit a tengerfenékről hiányzó hal és bálna csontvázak is bizonyítanak.
Vagyis a fosszilis leletanyag általános képe nem olyan, mint amit Darwin és követői megkívánnak. A szokatlan kövületek és az ősmaradványok felhalmozódására vonatkozó természetes magyarázatok hiánya (más, mint Noé bárkája) komoly kihívást jelent az evolúció számára.
A Kambriumi robbanás
Az egyetem utolsó nyári szünete alatt Darwin terepmunkát végzett Walesben Adam Sedgwick-kel. Ő az a Sedgwick, aki a kambriumi rendszert elnevezte (Wales ősi neve, Cambria után).*
Darwin teljesen tudatában volt annak, hogy van valami különös a Kambriummal kapcsolatban: számos olyan ősmaradványt találtak a kambriumi kőzetrétegben, amely úgy tűnt, hogy semmi primitívebb kövülethez nem kapcsolható az alatta lévő prekambriumi rétegekben. Ahogy a Fajok eredeté-ben megfogalmazta:
Arra a kérdésre, hogy miért nem találunk leleteket ezekből a rendkívül ősi időkből, nem tudok kielégítő választ adni. A legkiválóbb geológusok közül sokan, Sir R. Murchisonnal az élen, meg vannak győződve, hogy a legmélyebben fekvő sziluri (kambriumi) kőzetek szerves maradványaiban az élet hajnalát látjuk.36
Murchison a Teremtésre utalt és ez igen frusztrálhatta Darwint, amint néhány bekezdéssel később be is vallja:
A kérdés jelenleg megmagyarázatlan marad; és talán egy olyan valós magyarázat után kiált, amely cáfolja az itt bemutatott nézeteket.
Darwin végső megoldása menekülés volt a jövőbe, amikor a fosszilis lelet bizonyítékot szolgáltat arra, hogy neki van igaza és Murchison téved. Valójában a történeti tudomány egyes ironikus fordulataikor Murchison bizonyult hibásnak, amikor életet fedeztek fel a Prekambriumban, de Darwin is tévedett! Amíg egyes prekambriumi állatfajokat felfedeztek, úgy tűnik, hogy ezek is „berobbantak” a színre, és ebben semmi nem kapcsolja össze vagy magyarázza a kambriumi robbanást. Gould hozzáteszi:
…és a kambriumi robbanás problémája ugyanolyan masszív maradt, mint amilyen volt – ha nem még makacsabb, mivel a prekambriumi élet természetével kapcsolatban a zavar most már inkább a tudásunkon alapul, mintsem a tudatlanságunkon.37
Bizonyára igaza is volt, mivel az egyetlen ismert prekambriumi fauna (az Ediakara-fauna, lásd fentebb), amiről kiderült, hogy világszintű elterjedése volt, az evolucionisták szerint egy kudarcos kísérlet lehetett. Az ediakarai kövületeket nem tekintik a kambriumi és a későbbi életformák őseinek.
Miért maradtak a legegyszerűbb életformák, amelyek olyan gyorsan reprodukálódtak, egyedül olyan sokáig, továbbfejlődve a leghosszabb időn át valami olyanná, amely csak egy kicsivel komplexebb, csak hogy aztán a legváltozatosabb életformákká robbanjanak, amelyek valaha léteztek? Végtére is úgy tűnik, hogy elég furcsa és gyenge összefüggés van a darwini evolúció és a geológiai idő között. Mivel ez nagy kihívást jelent, a kambriumi robbanást az evolucionista körök többségében úgy tálalják, hogy az fogyasztható, viszont Darwin-szkeptikusok számára még nehezebben használható legyen. Ma leginkább úgy hivatkoznak rá, mint a kambriumi „lassú egybeolvadás”.
Ahogy egyre több ősmaradvány kerül napvilágra, ezeket a Kambriumon belül előre kijelölt pozícióba erőltetik bele, ezzel sokakat abban a hitben hagyva, hogy nem volt robbanás, és hogy a változás mértéke ezen időtartam alatt nem volt nagyobb, mint a földi élet legtöbb más periódusa alatt. Azonban a kék-zöld algák állítólag változatlanok maradtak 2,5 milliárd éven keresztül (nem beszélve az ezt követő fél milliárd évről, miután ma is morfológiailag változatlannak tűnnek), és ezután közel az összes törzs egy körülbelül 100 millió éves időkereten belül fejlődött ki. Figyelembe véve az evolúció újításainak ütemét, ez egy geológiai szempillantásnak felel meg.
Következetesen számba vesszük a tényeket és az olvasóra bízzuk, hogy maga vonja le a következtetéseket. Figyelmen kívül hagyjuk a fentebb tárgyalt rendhagyó kövületeket, különösképpen a prekambriumi rovarokat, annak érdekében, hogy a vita alapját kizárólag az elfogadott evolúciós adatok képezzék.
Ediakara után, a Kambrium legelején, egy másik furcsa állatfaj került a színre. A kelet-szibériai lelőhelye után elnevezett Tommoti fauna a meszes héjú állatok első megjelenése: brachiopodák, trilobiták, archaeociatidák (az első zátonyépítők, de nem korallok), puhatestűek, echinodermák és számos problémás teremtmény, amelynek nincs modern megfelelője. Nem sokkal később, a Kambrium középső szakaszában, az igazi „robbanás” akkor következett be, amikor a hihetetlenül sokszínű fauna hirtelen megjelent. Az elsőként felfedezett (1909), és azóta is leghíresebb helyszín a Burgess Pala a Brit-Kolumbiai Yoho Nemzeti Parkban. Később hasonló faunák kerültek elő a Grönlandi Sirius Passeten és a Kínai Yunan tartományban található Chengjiangban.
A Burgess Pala összesen 9 új törzset és 13 különleges ízeltlábút adott, amelyeket üledékekben se ezt megelőzően, se azóta nem találtak még.38 Ezek közül egyik sem származtatható (evolúciós értelemben) a korábbi fosszíliákból. Ami még sokkoló (a Darwinisták számára), hogy ezen új élőlények legtöbbjének olyan testfelépítése van, ami ma már nem létezik. Az ötszemű élőlények (Opabinia) és néhány, páratlan számú (7) szervpárral rendelkező faj azokban az időkben nem számítottak ritkaságnak. És később sem számítottak annak, mivel a leletek közül teljesen eltűntek. Napjainkban minden élőlény páros számú szervpárral rendelkezik. Ugyanez érvényes a szemre.
Ami igazán figyelemre méltó a Burgess Pala élővilágával kapcsolatban az az, hogy bár ez az összes eddig ismert állattörzs gyűjtőhelye, azóta nem jelent meg új törzs. A fosszilis leletek között nem találták még meg a testfelépítések ilyen szintű diverzitását. Gould ezt „tizedelésnek és változatosságnak” nevezte. Nézete szerint az élet úgy tűnik, hogy a tervezettség bőségével indult, ezek közül sok a későbbiek során eltűnt.
Ez feltehetően elegendő magyarázatot szolgáltat a valaha látott összes biodiverzitásra,39 de egyben a Darwini evolúció ellentétét is képezi. A természetes szelekciónak az egyszerűtől az összetett felé kellene haladnia. Ehelyett a legnagyobb evolúciós újítások hirtelen jelentek meg, előjelek nélkül, majd nem voltak további jelentős újítások, miközben az eredetiek közül sok kipusztult.
A neo-darwinista evolúcióbiológusok a Kambriumot megpróbálták a „szelekciós nyomás hiányá”-val magyarázni. Ez az értelmezés azon az elgondoláson alapul, hogy mivel gyakorlatilag az összes ökológiai állapot kialakulatlan volt, ezért majdnem minden, élet létrehozásával kapcsolatos kísérletezés (vagyis a mutációk furcsa és kevésbé furcsa kimenetelei) sikeresek lehettek, tekintve, hogy a komoly „versenyhelyzet” ekkor még a távoli jövőbe veszett. Ez természetesen a változásokra való gyors és folytonos képesség magas szintjét igényelhette, egy olyan lehetőséget, mely soha később nem megragadható. Gouldnak kételyei voltak ezzel kapcsolatban:
A kambriumi robbanás túl nagy, túl eltérő és túl egyedi volt. Egyszerűen nem tudom elfogadni, hogy ha az organizmusoknak mindig is megvolt a potenciáljuk ilyenfajta diverzifikációra – miközben csak az Alsó Kambrium különös ökológiája engedte meg valaha is ennek a megvalósulását – soha, egyetlen egyszer sem keletkezett új törzs a burgess-i idők óta.40
Gould és a „punctuated equilibrium” modell (hirtelen nagy evolúciós ugrások kicsi, csekély méretű populációkban, ellentétben a neo-darwinista hosszú idők alatti kis mutációkkal) más támogatói hivatkoznak az első komplex organizmusok sokkal nagyobb „genetikus rugalmasságára”, amely később elveszett a „genetikai rendszer elöregedésével”.41 Természetesen ezzel bármi megindokolható, amíg az evolúció nagy jelképét békén hagyják. Annak a lehetősége, hogy az összes törzset Isten hozta létre a Teremtés különleges eseménye során, és ez az Özönvíz során drasztikus módon tesztelésre is került, a világi tudósok teljes mértékben elvetik, még akkor is, ha ez egy egyszerűbb és teljesebb magyarázat. Amikor a geológiai és a paleontológiai tények láthatólag támogatják a magyarázatot, akkor ezeket a tudományos világ és a média csendben elhallgatja.
Fosszíliák, amiknek nem szabadna jelen lenni: a kivételes megőrződés esete
Sok éven át általános hiedelem volt, hogy mivel a fosszíliák ősidők óta halott élőlények megkövült maradványait mutatják, csak az organizmusok kemény részének van esélye fosszilizálódni hirtelen betemetődés nélkül. A kövületek többsége tengeri gerinctelenek vázaiból áll és azt gondolhatnánk, hogy a kövületek kis százalékát kitevő szárazföldi szervezetek után nem találunk millió éveket túlélő lágy részeket.
Ez magyarázatot adhat arra, hogy miért számított akkora meglepetésnek a Burgess Pala puhatestű tengeri élőlényeinek kivételes megőrződése. A fosszilizáció folyamatának kémiai bonyolultsága még ma is rejtély, ami a lassú és fokozatos folyamatokat illeti. Egyik felfedezés a másik után hökkenti meg a paleontológusokat és a biokémikusokat. Az igazán megdöbbentő, paradigmaváltó dolog a Dr. Mary Schweitzer által felfedezett puha, nem csontos szövet volt egy nem fosszilizálódott T. rex csontból.42 Az eredeti felfedezést további gondos munkálatok követték annak érdekében, hogy a kezdeti leleteket alátámasszák és kiegészítsék. Ma már számos dinoszaurusz egyedet találtak – különböző olyan fajokból, mint a teropodák, hadroszauruszok és a ceratopszidok43 – nem-fosszilizálódott szerves maradványokkal, amely egyben legújabban a dinoszaurusz DNS-hez szolgáltat bizonyítékot. Ha a lágyszövetek és a vérerek túlélhettek 65 millió évet vagy még többet, akkor a fosszilizáció teljes koncepciója újragondolásra szorul. Ez nem egy túl kellemes kilátás a paleontológusok számára. Mérhetetlen frusztráció jár ezzel együtt, mivel ez azt jelenti, hogy több száz év paleontológiai kutatásai és felfedezései során a megőrződött lágyszöveteket akaratlanul eldobhatták a csontokat körülvevő kőzetmátrix-szal együtt.
Nem meglepő, hogy az evolucionista világ kétségbeesetten próbálta kimagyarázni Schweitzer „veszélyes felfedezését” (ahogy a média nevezte) azt állítva többek között, hogy ez egyszerűen csak egy baktériumokból álló hártyaréteg („biofilm”), amely úgy néz ki, mintha lágy szövet lenne.44 Azonban Schweitzernek az ellenvetésekre vonatkozó válaszai olyan megalapozottak és meggyőzőek, amelyek a paleontológusokat meglepő állítások tételére vezették:
„Ezt mérföldkőnek nevezném”, mondja a paleontológus Hans Larsson a Montreáli McGill Egyetemről, aki nem volt részese a vizsgálatoknak. „A dinoszauruszok be fognak lépni a molekuláris biológia területére és valóban ki fogja lőni a paleontológiát a modern világba.”45
A lábasfejűek (mint a tintahal) lágyszövetű fosszíliáit már 1841-ben megtalálták, beleértve a köpenyt, a tintamirigyet, a szívókákkal és horgokkal felszerelt karokat.46 Ez az angliai Christian Malford és Trowbridge (Wiltshire) közelében található jura-kori Oxford Clay-ben történt a Nagy Nyugati Vasút építésének ideje alatt. A tudományos szakirodalomban szórványosan van megemlítve, mióta ez az elképesztő kövület-lelőhely 2000 után a tafonómiai vizsgálatok középpontjába került.47
A foszfatizációt, a szerves anyag foszfátásványokkal való helyreállítását, ma úgy tekintik, mint a kivételes fosszilizáció okát.48 2007-ben ezek a kövületek médiaszenzációvá váltak, mivel a Belemnotheutis antiquus ép állapotban megmaradt zsákocskájából származó tintát ammóniával „aktiválták újra”, ami a paleontológusok számára segítség volt a tintahal alakjának felvázolásához!49 Valójában ez egy régi praktika, azonban ezen fosszíliák forráshelyszíne száz vagy még több évig ismeretlen volt. Ami igazán figyelemreméltó és mindmáig nem teljesen érthető, hogy a foszfatizáció hogyan zajlódhatott le ilyen gyorsan annak érdekében, hogy gátolja a lágyszövetek és a könnyen bomló melanin foszlását, annak az anyagét, amely a sötét tinta színezőanyagát alkotja.
Németországban a kivételesen megőrződött fosszíliák helyszínei kellőképpen gyakoriak ahhoz, hogy saját, német eredetű elnevezésük legyen: Lagerstätten (kövülettelér). Ezt a szót használják ma hasonló helyszínekre szerte a világon. Ida50, az eocénbeli főemlős, akinek az állítólagos őse nem túl sok évvel ezelőtt megőrjítette51 a médiát (és sok tudóst is a média szenzációhajhászása miatt) a németországi Messel-gödör lelőhelyről származik.
A világ összes lelőhelyeinek gyakorisága és kiterjedtsége a kivételes megőrződést inkább általános szabállyá teszi, mintsem kivétellé. Ez a fosszilis leletek darwini megközelítésének egy alapvetően feloldhatatlan ellentmondása. És egyben a ‘fiatal föld’ nézetének egy teljesen ésszerűvé tétele.

KÉP:
A Darwinius masillae („Ida”) egy jól megőrződött fosszíliája, ami körül felfokozott médiakampány zajlott. Ida közel azonos volt a mai lemurokkal.50
Átmeneti fosszíliák
Miután számba vettük a fosszilis leletekkel együtt járó számos evolúciós Achilles sarkot, beleértve a főbb taxonómiai csoportok és több kívül eső kövület hirtelen megjelenését, vizsgáljunk meg néhányat a legfrissebb és legerősebb állítások közül, melyekkel az evolúciós átmenetek mellett érvelnek.
Elsőként látnunk kell, hogy az evolúciós átmenet bizonyítékaként használt fosszíliák túlnyomó többsége csak kisarányú varianciát mutat, amely egy alapfajon belüli variációként írható le. Ez beleillik abba a közismert kreacionista elképzelésbe, miszerint Isten sokféle, egymástól jól elkülöníthető állatfajt teremtett, mindegyiket gazdag genetikai változatossággal és a mérsékelt változás képességével a különféle környezeti feltételekhez való alkalmazkodás és a túlélés érdekében. Ez a változás magában foglalhatja a fiziológiai alkalmazkodást a megtervezett genetikus variáción végbemenő szelekción keresztül, valamint a mutációk okozta variációt is.
Emiatt, amikor egy bizonyos leszármazási vonalon belül lezajló apró változásokat vizsgálunk, ez nem egy perdöntő bizonyíték. Így nem használható fel bizonyítékként egyik oldal mellett sem. Ez az egyik ok, amiért a bibliai kreacionisták rendszeresen amellett próbálnak érvelni, hogy a nagy változásokat mutató dolgok (és az erre vonatkozó bizonyítékok hiánya), az élet eredete, és az információtartalom eredete ennek az életnek a velejárója. Ezek jelentősen nagy változások és az evolúciótannak egy jobb tesztelését teszik lehetővé.
A kihalt állatok csontjainak, nyomainak különálló evolúciós kategóriákba való helyezésének nehézségeit figyelembe véve Maciej Henneberg professzor egy ízben ezt írta:
Nincs pontos módszer annak megállapítására, hogy vajon Julius Caesar és Diana hercegnő azonos fajhoz, a Homo sapiens-hez tartoztak -e.52
Megkérdezheti valaki: „Hogyan határozható meg egy ‘átmeneti fosszília’?” Az evolucionisták válasza egyszerű: „Az átmeneti fosszília egy olyan organizmus, amely közös jellemzőkben osztozik mind az elődeivel, mind az utódaival.” Ezután a következő kérdés ez lenne: „Honnan tudhatja valaki, melyek az elődök, melyek az utódok?” A válasz így szól: „Az előd korábban élt (és korábbra datált rétegekben található, mint az átmeneti fosszíliák) és több primitív jellemzővel bír. Az utód később élt (fiatalabbra datált rétegekben, mint az átmeneti fosszíliák) és fejlettebb jellemzőkkel rendelkezett/rendelkezik.”
Nos, ez lehet a helyzet minden adott időpillanatban, de a későbbi felfedezések bármely fosszília átmeneti státuszát módosíthatják. A paleontológia történetében ez sokszor előfordult. Ráadásul a fosszília-típusok közötti határok rendkívül rugalmasak, ahogy később látni fogjuk. Hogy még tovább bonyolítsuk a helyzetet, ki állapítja meg, hogy a fosszília hol foglal helyet az evolúciós létrán? A kövületek fel és le is mozoghatnak a felfedezéseknek és az elméleteknek megfelelően. Természetesen ez jobbára szakmai titok, a paleontológiai adatok átlagfogyasztója nincs tisztában vele.
Henneberg fentebbi megfigyelése kijózanító; ha nem tudjuk letesztelni, hogy két ember, akiket 2000 év választ el, vajon azonos faj tagjai-e, hogyan tudnánk megtenni olyan élőlényekkel, amelyeket állítólag több millió év választ el? A valóságban ez egy hatalmas probléma, de az evolucionista paleontológusok ezt vidáman elbagatellizálják, és minden további nélkül úgy rendezik el az egymástól több száz millió évre lévő kövületeket, hogy beleilleszkedjenek az evolúciós sorba és narratívába (végül egy kladogramba, ahogy korábban szó volt róla).
Az Archaeopteryx és dinoszaurusz-madár átmenet
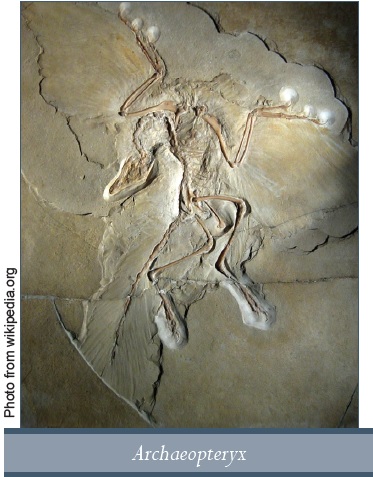 Az Archaeopteryx az első olyan fosszília, amelyről azt állították, hogy a dinoszauruszok és a madarak közötti átmeneti forma. Később ősi madár lett, de 2011-ben egy másik kínai fosszília, a Xiaotingia felfedezése után az Archaeopteryx-et átsorolták egy therapodához hasonló dinoszauruszhoz, a Deinonychus-hoz53, mielőtt egy másik kutatócsoport áthelyezte volna, kifogásolva az első megállapítását. Ami magát a Deinonychus-t illeti, szintén átment egy érdekes átalakuláson, amint ezt a rekonstrukciója melletti tábla összegez egy tollas dinoszaurusz kiállításon:
Az Archaeopteryx az első olyan fosszília, amelyről azt állították, hogy a dinoszauruszok és a madarak közötti átmeneti forma. Később ősi madár lett, de 2011-ben egy másik kínai fosszília, a Xiaotingia felfedezése után az Archaeopteryx-et átsorolták egy therapodához hasonló dinoszauruszhoz, a Deinonychus-hoz53, mielőtt egy másik kutatócsoport áthelyezte volna, kifogásolva az első megállapítását. Ami magát a Deinonychus-t illeti, szintén átment egy érdekes átalakuláson, amint ezt a rekonstrukciója melletti tábla összegez egy tollas dinoszaurusz kiállításon:
Ezek a szobrok eredetileg pikkelyes bőrrel lettek elkészítve 1986 és 1989 között más dinoszauruszokból származó bőrlenyomatok alapján. Amikor a Deinonychus-t először 1969-ben megtalálták, úgy gondolták, hogy egy madárszerű dinoszaurusz és a madarak egy lehetséges őse. Ma már tudvalevő, hogy a Deinonychus-nak repülő ősei voltak – dromeoszauruszok – amely inkább a röpképtelen madarak egy formájává teszi őt, mintsem dinoszaurusszá. A Deinonychus-t a kínai repülő dromeoszaurusz fosszília felfedezése után megtalálva a tudósok nem tekinthettek rá pikkelyes dinoszauruszként, hanem egy olyan madárként, amely elvesztette a repülésre való képességét.54
Ez olyan messze van az empirikus tudománytól, amennyire csak lehet, nem igaz? Hogyan lehet egy dinoszaurusz csontvázat (és bőséges Deinonychus-fosszília áll rendelkezésre) először dinoszaurusznak minősíteni, aztán egy röpképtelen madárnak? Rengeteg anatómiai részlet őrződött meg a csontkövületekben, olyan sok, hogy a madár elválasztásának a dinoszaurusztól sziklaszilárdnak kell lenni (szándékos szójáték). Azonban mégis az történt, hogy a madarak és a dinoszauruszok közötti határ szándékosan össze lett mosva azért, hogy gyakorlatilag bármely jövőbeli felfedezés „újratervezhető” legyen.

KÉP: Bár a Sinosauropteryx volt a dinó-madár átmenet eredeti „kirakatbabája”, azok a rostok, amelyeket tollakként írtak le, elég egyértelműen bőr alattiak.
A legegyszerűbb magyarázat (az Occam borotvájának megfelelően) nem része az evolucionista biológusok gondolkodásának: minden tollas fosszília madárfosszília és minden madárszerű, tollnélküli fosszília a dinoszauruszokhoz tartozik. Más szavakkal, a tollak megléte lenne a döntő követelménye a madarak dinoszauruszoktól való elválasztásának. Valójában egy másik híres „tollas” dinoszaurusznak, a Sinosauropteryx-nek egy friss vizsgálata mutatta be nagyon világosan, hogy a „tollak” egy sűrű, bőr alatti réteg részei voltak. A tanulmány szerzői ezt mondták:
Meglehetősen szokatlan módon Currie és Chen (2001) csak a rostszálas szöveteket ismertette a NIGP 127587-ben (mint őstollakat) annak ellenére, hogy a bőrszövet központi jelentőségű és figyelmen kívül hagyja, hogy ez magába foglalja a külső szövet nagyobbik részét, a fejtől a farokig összefüggő rétegként (1. és 6. ábrák). Ez pedig szinte félreérthetetlen. Az itt bemutatott leírás azt mutatja, hogy a rostszálas struktúrák belső védőrostok voltak …55
Vannak más problémák (melegvérű kontra hidegvérű, hüllőtüdő vs madártüdő), amelyeket szintén figyelembe kell venni. A dinoszauruszok hüllők voltak, de sajátos jellemzőikkel egy külön hüllőcsoportot alkottak. Ők szintén kihaltak, miközben a többi hüllőcsoport nagy része a dinoszauruszokkal együtt temetődött el, mégis a mai napig élnek. Lehetnek bizonyos jellemzőik, amelyek a mai madarakéhoz hasonlóak, de ez nem elegendő ahhoz, hogy bemutassuk a dinoszaurusztól a madárig vezető evolúciót.
A másik lehetőség, hogy a dinoszauruszoknak valóban voltak tollaik. Hogy a madarak fejlődtek-e ki a dinoszauruszokból, az egy más kérdés. Miközben igaz, hogy az evolucionisták a Theropoda dinoszauruszok közé helyezték el a madarakat, a kreacionistáknak nincs előzetes állásfoglalásuk a tollas ill. tollnélküli dinoszauruszokról vagy arról, hogy a madarak a dinoszauruszokkal összecsoportosulhatnak-e egy egymásba illesztett hierarchiában. Ez igaz, mivel a Biblia nem tesz konkrét előrejelzést arról, hogy a madarak hová csoportosíthatók, és nem is említi a dinoszauruszokat (név szerint).
Habár a taxonómia atyja (Linné) kreacionista volt, ez nem jelenti azt, hogy az osztályozása szent és sérthetetlen, sőt az is igaz, hogy eltérő csoportok közös jellemzőik alapján egy kalap alá vehetők (pl. az ember a majmokkal és a selyemmajmokkal együtt az emlősök közé tartozik). Mégis a madarak dinoszauruszokból származó evolúciója egy evolúciós „hadszíntér”.
Tiktaalik
 Amikor a Tiktaalik roseae-t 2004-ben az Észak-Kanadai Ellesmere56 szigeten felfedezték, rögtön az úgynevezett átmeneti fosszíliák szűk klubjához csatlakozott, nagy médiafelhajtás közepette. Úgy tűnt, hogy a kreacionisták evolúció elleni érvelése a végét járja, mivel ez a hal-lábú „bizonyosan” a halaktól a négylábú állatok (tetrapodák) felé tartó átmeneti forma volt. Ezt a közép-devoni időszakra datálták (383 millió évvel ezelőtt), az első ismert tetrapodák elé.
Amikor a Tiktaalik roseae-t 2004-ben az Észak-Kanadai Ellesmere56 szigeten felfedezték, rögtön az úgynevezett átmeneti fosszíliák szűk klubjához csatlakozott, nagy médiafelhajtás közepette. Úgy tűnt, hogy a kreacionisták evolúció elleni érvelése a végét járja, mivel ez a hal-lábú „bizonyosan” a halaktól a négylábú állatok (tetrapodák) felé tartó átmeneti forma volt. Ezt a közép-devoni időszakra datálták (383 millió évvel ezelőtt), az első ismert tetrapodák elé.
Mégis mindössze 6 év kellett ahhoz, hogy ez is csatlakozzon a Deinonychus, a Sinosauropteryx és sok más átmeneti státuszban lévő jelölt sorsához, mivel valódi tetrapoda lábnyomokat találtak Lengyelországban és 10 millió évvel a Tiktaalik elé datálták!57

KÉP: Mészkőtábla megkövesedett lábnyomokkal Lengyelországban.
Az evolucionisták saját érvelésével a szegény hal-lábút egyszerű maradványként kell kezelnünk, feltételezett átmeneti jellemzőinek legalább 400 millió évvel ezelőtt kellett kifejlődniük (időben azt a tetrapodát megelőzve, amely a lábnyomokat produkálta, és amely teremtmény mindmáig felfedezetlen).
Az átmeneti fosszíliák leleteken belüli hiánya természetesen önmagában nem érv a létezésük ellen. Tehát az az evolúciós magyarázat, hogy a fosszilis lelet csak az adott időpillanatot őrzi meg, az átmeneti formák pedig a különböző időszakokról tanúskodnak, nem teljesen abszurd. Azonban képtelenség és nyíltan tudományellenes azt állítani, hogy a hiány nem használható fel érvként az egész átmeneti formákkal kapcsolatos koncepcióval szemben, különösen mivel az a történet, amit a fosszilis leletek mutatnak messze nem kielégítő az evolucionista paleontológusok számára:
Nem csoda, hogy a paleontológusok már régóta menekülnek az evolúciótól. Úgy tűnik ugyanis, hogy soha nem történt meg. Kitartóan felhalmozódva a sziklafalak cikkcakkokat, apró rezgéseket produkálnak és a nagyon esetleges változások millió évek alatt túl lassú ütemben halmozódnak fel, hogy valódi magyarázatul szolgáljanak minden olyan bámulatos változásra, amely az evolúciós történelem alatt lezajlott. Amikor látjuk az evolúciós újítások megjelenését, gyakran robbanásszerűen bukkannak fel, és gyakran erős bizonyíték nélkül arra vonatkozóan, hogy az organizmusok nem evolválódtak máshol! Az evolúció többé nem zajlódhat le más helyen. Mégis a fosszilis lelet sok tanácstalan paleontológust ilyen módon késztetett arra, hogy tanuljon valamit az evolúcióról.58
Paradox módon az a kevés átmeneti forma, amiről Darwin tudott, nem volt használatban a Scopes Trial (1925) idején, és az a győzelmi lista, amit az 1960-as, 70-es években láttunk, mára gyorsan lecsökkent egy roppant kevés számúra, és a fennmaradtak egyike sem mentes a súlyos problémáktól.
Természetesen a népszerű weboldalak az átmeneti formák hosszú listáját vázolják fel, de nem mutatnak rá arra a hatalmas különbségre, ami az azonos fajon belüli variációk (vagyis olyan alapcsoport, mint a ‘család’) és az egyik fajtából a másikba való átmenet között húzódik. Ki állítaná azt, hogy egy átlagos és egy apró méret közötti Schnauzer kutyafajta átmeneti forma lenne? Ők mindannyian a Schnauzer fajtához tartoznak. A fosszilis leletek között rengeteg variációt látunk bizonyos kereteken belül, és nagyon kevés köztes forma van a nagyobb állat- és növénycsoportok között. Ekképpen az evolúciós történelem legnagyobb előrelépései azok, amelyek az innováció legmagasabb fokát igényelték, és amelyek megszámlálhatatlanul sok kísérleti evolúciós prototípust fogyasztottak el, ezek vannak a legkevesebb számú köztes forma által áthidalva. A valóság pontosan az ellenkezője, mint amit Darwin előrejelzett!
Puijila
2009-ben a Puijila darwini-nek elnevezett fosszíliát úgy jelentették be, mint a legutolsó ‘összekötő kapocs’.59 Ez volt az első olyan bizonyíték, amelyet az úszólábúaknak (fókák, oroszlánfókák, rozmárok) a szárazföldi emlősökből származó evolúciójának tulajdonítottak. Richard Dawkins ujjongva kiáltott fel:
Puijila találóan fogja közre az úszólábúak szárazföldi és vízi őseik között lévő szakadékot. Ez egy újabb örömteli adalék az ‘összekötő kapcsok’ egyre növekvő listájához, amelyek nem hiányzóak többé.60

E Függelék: Puijila csontváz
Úszólábú vagy vidra?
Habár az evolúciót támogató tudósok “sétáló fókaként”3 vagy fókafejű vidraként22 utalnak a Puijilára, csontvázának felépítése nagyon hasonlít a folyami vidráéra, amint a lenti fotó is mutatja.
Puijila csontváz
Folyami vidra csontváz
Dr. Werner az Evolution: the Grand Experiment (Evolúció: a nagyszabású kísérlet) című könyvének 2. kiadásából származó E függelék egyik oldala bemutatja a feltételezett átmeneti forma csontvázát (fent) és az Észak-Amerikai folyami vidráét (lent). Azon kívül, hogy a Puijila lúdtalpas, nehéz további különbséget észrevenni.
Van egy komoly probléma a Puijila evolúciós magyarázatával, habár csontvázát tekintve majdnem azonos az Észak-Amerikai folyami vidrával, a Lontra canadensis-sel.61 Ilyen módon nemcsak a Puijila nem egy átmeneti fosszília, hanem ez egy ma élő faj maradványa, tehát az úszólábúaknak továbbra sincs evolúciós történetük. Ez annak a ténynek ellenére igaz, hogy máig több, mint 15.000 úszólábú fosszíliát fedeztek fel.62
Bálnák
A bálnák evolúciója manapság felkapott témának számít. Ősi bálnák és bálnaszerű élőlények egész sora, amely egy pár évtizeddel ezelőtt még ismeretlen volt, ma már elérhető, olyan módon elrendezve, amely egy evolúciós leszármazási sornak megfelel. A bálnák evolúciója három fosszílián nyugszik, amely összeköti az ‘első’ valódi bálnákat (Basilosauridok) a szárazföldi állatokkal: a Pakicetus-szal, az Ambulocetus-szal és a Rodhocetus-szal, de mind a három megbukott, amint arra Dr. Carl Werner, az Evolution: the Grand Experiment szerzője rávilágított.63
A Pakicetus-szal kapcsolatban Dr. Philip Gingerich elképzelt egy koponyatöredéket, amely egy bálnaszerű élőlényé lehetne, ez szerepelt a Science magazin címlapján 1983-ban. Amikor 2001-ben a Pakicetus maradványainak többi részét is publikálták, akkor bizonyítást nyert, hogy egy szárazföldi állatról van szó.64 Egy 2009-es National Geographic dokumentumfilmben Dr. Gingerich továbbra is azt állította, hogy a Pakicetus-t az fülcsontja alapján a bálnák közé kellene sorolni. Azonban a fülcsont egyáltalán nem olyan, mint a bálnáé, amelynek ujjszerű (S-alakú) kinövése van. Nem, ez lemezszerű, olyan, mint a párosujjú patásokként ismert szárazföldi emlősöké.
Dr. Hans Thewissen azt állította, hogy az Ambulocetus nyolc jellemzője egy ősi bálnáé. Dr. Werner feljegyezte Dr. Thewissen abbéli beismerését, hogy a bálnaősre vonatkozó kulcsbizonyíték, a fülcsont S-alakú kinövése valójában nem másé, mint egy bálnáé volt. Dr. Werner kijelenti: „Mind a nyolc jellemző, amiről [ő] bálnatulajdonságként számol be, zavarba ejtően nem bálna jellemzők.”
A Rodhocetus-ról azt állították, hogy egy tengeri emlős, amely úszólábat és bálnaszerű farkat növesztett – a bálnává válás irányába. Azonban nem volt fosszilis bizonyíték sem a farokra, sem az uszonyokra.65 Amikor erről a kérdésről faggatták, Gingerich beismerte, hogy a későbbi leletek alapján ezentúl nem hiszi, hogy a Rodhocetus-nak horgas farka vagy uszonya lett volna.64 Tehát a bálna evolúciójának átmeneti sorába tartozó kulcsfontosságú állat nem az, amiről azt állították, hogy az.

KÉP: Rodhocetus a Michigani Egyetem Természettudományi Múzeumában, USA. Az eredeti fosszilis lelet kiegészítve a múzeumi illusztráción. Dr. Carl Werner, Evolution: the Grand Experiment (Evolúció: a nagy kísérlet) Vol. 1 DVD. A piros x-ek jelölik a képzelőerő felhasználását az illusztráción.
Sok más probléma van a bálnák evolúciójával, habár a történetük logikusan le van vezetve. Az evolucionisták még azon sem egyeznek meg, hogy melyik szárazföldi állatból alakultak ki a bálnák. A fogak fosszilis hasonlóságainak alapján egyes paleontológusok a hiénaszerű állatokat (Pachyaena) részesítették előnyben, míg mások a macskaszerű állatokat (Sinonyx). Azonban friss DNS-összehasonlító vizsgálatok alapján a molekuláris biológusok úgy döntöttek, hogy a víziló volt a bálnák legközelebbi őse. De nagy problémák adódnak a vízilónak bálnává való átalakításával (például egy szárazföldi állat hogyan szerzi meg azt a képességet, hogy életet adjon kicsinyének és gondozni tudja azt víz alatt!).
Élő kövületek
A világ evolúciós történetéből – legalábbis annak média- ill. könyvverziójából – az a benyomásunk alakulhat ki, hogy az evolúciós leszármazás a természetes szelekción átszűrve a teljes univerzumnak egy megállíthatatlan és mindenütt jelenlévő jellemzője, amelynek az élővilág csak egy parányi része. Ezért meglepetésként éri a lelkes tudóst, hogy bőséges az evolúciós pangás eseteinek száma (amely minden meghatározás szerint egy oximoron). A tény az, hogy megszámlálhatatlan olyan élőlény létezik, amely nem, vagy a különösen hosszú geológiai idő alatt csak nagyon kis mértékben változott.
Erre a nyilvánvaló összeférhetetlenségre a sztenderd válasz az, hogy evolúció csak akkor zajlik, ha szükséges (vagyis “Az élőlénynek nem volt szüksége arra, hogy evolválódjon, mivel már tökéletesen alkalmazkodott a környezetéhez.”) Másképp fogalmazva, a természetes szelekció csak azokat a módosulásokat válogatja ki, amelyek nem a legjobban adaptálódtak egy adott környezethez. Habár a környezet nem változik, a már kiválogatódott legjobb módosulások vagy variációk lesznek azok, amelyek idővel továbbélnek. Ezzel a redukcionista megoldással azonban komoly problémák adódnak, ideértve azt a tényt, hogy az élő kövületek állítólag túlélték a legnagyobb globális katasztrófákat (pl. a késő permi tömeges kihalást, amely állítólag 252 millió évvel ezelőtt történt és a később élő fajok többségét kipusztította, valamint a 65 millió évvel ezelőtti, Kréta-harmadkor környéki meteorbecsapódást, amely eltörölte a dinoszauruszokat). Ezeket az eseményeket drámai klímaváltozásokkal hozzák összefüggésbe az evolúciós modellekben. Emiatt csak rendkívül kicsi azoknak a mikrokörnyezeteknek a száma, amikről úgy vélik, hogy változatlanok maradtak. Hogyan tudott akkor bármilyen élőlény “tökéletesen alkalmazkodva” megmaradni? Ugyanakkor, hogyan tud egy faj azonos maradni miközben más fajok evolválódnak, és versengenek velük és/vagy megeszik őket? Maguk az evolucionisták vallották be a problémát:
De a pangást, mint az élet történetének egy olyan tényezőjét, amit számításba kellene venni, egyszerűen száműzték az evolúcióbiológiából. A pangást egészen addig nem vették figyelembe, míg Gould és jómagam rámutattunk, hogy egy ilyen értelemben vett stabilitás az élet történetének egy olyan valós aspektusa, amellyel szembe kell nézni – és hogy valójában nem jelent alapvető fenyegetést az evolúció alaptézisére. Ez volt Darwin problémája: ahhoz, hogy megalapozza az evolúciótan elfogadhatóságát, úgy érezte, alá kell ásnia korábbi … alaptételét a fajok változatlanságáról. Darwin számára a sztázis egy zavaró kellemetlenség volt.66
Gould és Eldredge hitte, hogy a punctuated equilibrium – az evolúció hirtelen nagy ugrásai hosszú pangási időszakok után – volt az evolúció valódi sémája. Azonban ők még csak meg se próbáltak arra magyarázatot adni, hogy honnan származhatott a genetikai információnak ilyen masszív gyarapodása. Így, miután Gould elhunyt, a punctuated equilibrium vesztett népszerűségéből, és a gradualista neo-darwinizmus vált újra a tudományos élet vezető dogmájává. Ennek következtében az evolúciós pangás problémája egyéb más zavaró kérdések sorsát követte, így ezek különböző nyakatekert görög szavakkal és kifejezésekkel lettek elkendőzve.
Sztromatolitok
A Föld „legidősebb” kövületei a sztromatolitok (állítólag 3,5 milliárd évesek), amelyeket a kék-zöld alga (cianobaktérium) kolóniák maradványainak tartanak. Amíg kék-zöld algák még ma is élnek, meglepetésként hatott, hogy e mikroorganizmusok kolóniában élő formái, melyeket a fosszilis leletekből, sztromatolit néven ismerünk, szintén élnek, láthatóan változatlan formában! A hasonló eseteket népszerű nevén „élő kövületeknek” hívják, mert a fosszíliát az élő állatok előtt találták. Nyelvészeti szempontból ez a kifejezés ugyanolyan abszurd, mint a “nem-halott”.

KÉP:
A bahamai és ausztráliai élő sztromatolitok tanulmányozása azt mutatja, hogy fosszilis formáik megegyeznek a ma élő változatokkal.*
Awramik, S.M., Respect for stromatolites (Tisztelet a sztromatolitoknak), Nature 441:700–701., 3. ábra
Nem kétséges, hogy a sztromatolitok jelentik az evolúciós pangás lényegét, mégis rengeteg probléma van egy ilyen interpretációval. Először is mivel a neo-darwini dogma úgy tartja, hogy a mutációk az evolúció motorjai, az ebben való hit a logikát a megismerésen túlra feszíti, vagyis hogy a telepes prokarióták mutációi 3,5 milliárd év alatt sem voltak rájuk nagyobb hatással! Mindenesetre a neo-darwinizmus szerint minél egyszerűbb az élőlény annál gyorsabb az evolúciója, mivel minden mutáció jelentős és ezeket az egyszerűbb élőlények rövid idő alatt továbbadják a következő generációnak. Ha csak figyelembe vesszük a sztromatoliokban/kék-zöld algákban az állítólag 3,5 milliárd év alatt végbemenő véletlenszerű mutációk számát, a sztázisuk szembemegy a logikával.
Másodszor, amint fentebb említettük, a bolygó evolúciós története szerint a környezetre sok masszív változás volt hatással. És mivel ma is találnak élő sztromatolitokat a világ ellentétes oldalain, Ausztráliában és a Bahamákon, biztosan következtethetünk arra, hogy a geológiai idő során ezek komoly környezeti kihívásoknak voltak kitéve – az feltételezett evolúciót még jobban elősegítve. Mégsem ez történt.
Tőrfarkú rák
A Limulus nemzetség “legkorábbi” ismert tagja a kanadai közép- és észak-Manitobából származik, kövületei a késő ordovíciumi üledékekből kerültek felszínre, 445 millió éves állítólagos korral – amely a többsejtű organizmusok evolúciós történelmét nézve nagyon korai. Azon túl, hogy kisebb – miközben tudjuk, hogy a méret egyszerűen csak a megkövült egyed életkorát tükrözi –, maga a fosszília ránézésre megegyezik a mai tőrfarkú rákkal. A felfedezők szavai sokatmondóak:
Ha megértjük, hogy a tőrfarkú rákok hogyan alkalmazkodtak már nagyon korán ehhez az ökológiai niche-hez, és minden nehézség ellenére megmaradtak, segít bepillantani abba, hogy az óceáni ill. partvonal menti ökoszisztémák hogyan fejlődtek a hosszú idő alatt.67
És ismét, tekintetbe véve (az evolucionisták szerint) az Ordovícium óta lezajlott környezeti változások volumenét, ennek a teremtménynek a változatlan fennmaradása egy csoda, és kihívás elé állítja a gradualista neo-darwinizmust. Azonban ők ezt nem látják a hosszú korszakokon alapuló világnézetükkel ellentétesnek, mivel feltételek nélkül hisznek benne. Az ő világnézetükön belül minden rendellenesség egy “izgalmas” felfedezéssé válik.
A bojtosúszós maradványhal
Az 1839-ben, Ausztráliában történt első felfedezésük alapján a bojtosúszós maradványhalakról úgy hitték, hogy egy átmeneti evolúciós forma a halak és a négylábú szárazföldi állatok között. Az izmosúszójú halak e csoportjának küvületeit a Devontól a késői Krétáig datált üledékekben találták meg, amely időszakról úgy gondolják, hogy ekkor halt ki. Izmos uszonyaira úgy tekintettek, mint a lábak előjeleire.
1938-ban egy élő bojtosúszójú halat fogtak Dél-Afrika északi partjainál. Ez hatalmas megdöbbenést okozott nemcsak azért, mert ez a hal élt és egészséges volt, hanem mert lényegében 400 millió évig változatlan maradt, beleértve a 65 millió évvel ezelőtti “kihalását”. Azóta sok helyen találtak élő bojtosúszójú halakat, és le is filmezték őket természetes élőhelyükön a Comore-szigetek közelében.
Kiderült, hogy ezek a mélytengeri halak semmilyen módon nem fejlesztettek ki lábakat. Azon a tényen kívül, hogy a természetes élőhelyük messze esik bármilyen tengerparttól, izmos uszonyaikat arra használták, hogy ügyesen elnavigáljanak víz alatti élőhelyükön, valamint hogy fürgén kövessék az áramlatokat, vagy ha azt akarták, egy helyben maradtak. Uszonyaikat … úszásra használták! A nemzetség más fajait az 1990-es években találták meg, amelyek csemegének bizonyultak az Indonéz halpiacokon. Viszlát összekötő kapocs, hello új (és fincsi) élő kövület!
Skorpiók és szitakötők
A legkorábbi, állítólag több mint 360 millió éves szárazföldi skorpió-fosszíliát Kínában68 találták felső-devoni üledékekben. Habár a lelet hiányos, nagyon hasonlít a mai skorpiókra. Egy hatalmas skorpiót, a Pulmonoscorpius kirktonensis-t Skóciában69 fedeztek fel, fiatalabb széntartalmú üledékekben, amely az evolúciós számítások alapján közel 300 millió éves. Anatómiai jellegzetességeinek többsége megegyezik a modern skorpióéval, az egyetlen nagyobb különbség a méret. 70 cm hosszúra is megnőtt, miközben a legnagyobb élő skorpiók 20 cm körüliek.

Érdekes, hogy az ízeltlábúak (ahova a skorpiók is tartoznak) között az óriásnövésnek más példái is vannak a széntartalmú üledékekben: a Meganeura egy olyan szitakötő, amelynek a szárnyfesztávolsága 75 cm és az Arthropleura – a legalább 2 m hosszú százlábú. Erről az óriásnövekedésről sokan helytelenül azt gondolják, hogy a Karbon időszak magasabb atmoszferikus oxigén-koncentrációja okozta. Mivel a skorpiók és a szitakötők alapfelépítése láthatóan változatlanul megőrződött 320 millió éven át, élő kövületként is osztályozhatóak.
A Wollemi fenyő
1994-ben egy felettébb meglepő felfedezés egy új növényt adott hozzá az élő kövületek listájához. Az ausztráliai Új-Dél Wales területén, a Blue Mountain (Kék-hegy) egy mély hegyszorosában, Sydney nyugati részén, mintegy 5.000 m2-es területen, egy Wollemi fenyő-ligetet fedeztek fel. A fa valójában nem fenyő, hanem a chilei araukária fa egy rokona, és addig csupán kövületként volt ismert a Jura időszakból. Amikor először nyilvánosságra került, a felfedezés olyan szenzációt gerjesztett a média- és tudósvilágban, hogy azt mondták: „ez olyan, mintha dinoszauruszt találnál a hátsó udvarodon”. Néhányan óvatosságra intettek mondván, hogy ez egy „új nemzetség” és túl korai lenne élő kövületnek hívni.70 Azonban azt senki nem tette szóvá, hogy a lelet összeegyeztethetetlen az evolúciótannal. Ma már ennek a növénynek állandó helye van az élő kövületek listáján. Miközben széles körben termesztik és árusítják őket kertészetekben, habár a népszerűsítésükre irányuló, kormányzat által támogatott projekt a gyenge eladási számok miatt leállt.71
Élő medúza
Az “élő kövület” egyik legjobb példája a Utah állambeli sziklák között található. Ez a medúza nemcsak szépen megőrződött, hanem mivel nagyon hasonló állatok léteznek ma is, ezért úgy tartják számon, hogy ez a csoport 505 millió év alatt lényegében változatlan maradt.72 Ezt a félmilliárd évet a Kambrium éra közepére teszik, amikor a többsejtű élet először kialakult. Ily módon ez a medúza a többsejtű földi élet teljes evolúciós időszakán át változatlan maradt, amíg egyes rokonai emberré fejlődtek tovább! Látja-e a kedves olvasó, hogy ha az evolúció egyrészt állítja, hogy képes megmagyarázni az időben lezajló radikális változásokat, másrészt azt is állítja, hogy magyarázatul szolgál a nagyfokú pangásra, akkor nem lesz olyan bizonyíték, amelyet be lehetne vetni az evolúciós teória ellen? Valójában ez inkább filozófia, mint tudomány.

KÉP:
Egy kivételesen épen megőrződött, meglehetősen modern kinézetű medúza, amely állítólag félmilliárd éves.
Cartwright, P., Halgedahl, S.L., Hendricks, J.R., Jarrard, R.D., Marques, A.C., Exceptionally preserved jellyfishes from the Middle Cambrian (Kivételes módon megőrződött közép-kambriumi medúzák), Public Library of Science ONE 2(10):e1121.doi:10.1371/journal.pone.0001121, October 2007.
Megkövült nyomok
Az állati tevékenyég nyomai (lábnyomok, farknyomok, fészkek, odúk stb.) már régóta ismertek a fosszilis leletekből. A tudományban ichnite-ként ismert szóból (a „lábnyom” görög megfelelője) eredeztetik a paleontológia új ágát, az ichnológiát. Az összes különböző nyom, élőhely stb. egy frappáns latin névvel lett jelölve a kettős nevezéktan szerint, annak ellenére, hogy a hozzájuk tartozó állatok ismeretlenek voltak. Ez nagy mennyiségű név-ismétlődést idézett elő. Az esetek túlnyomó többségében egy ichnológiai elemnek, bármi is legyen az elnevezése, van egy ismeretlen forrása, amely azt jelenti, hogy egyazon rétegben találhatunk lábnyomokat, amelyek egy ismeretlen élőlényhez tartoznak, valamint egy ismert teremtmény megkövült maradványait, amely nem köthető egyértelműen a nyomhoz.73 Más esetekben a későbbi felfedezések összekötötték a nyomot az ismert fosszíliával. Például az óriás százlábú, a (fentebb említett) Arthropleura nyomaihoz lett hozzárendelve egy ichnotaxon név, a Diplichnites cuithensis.74 A százlábú nyomához ez egy frappáns név! Ez kibővíthető sok egyéb bonyolító elemmel. Például azzal a ténnyel, hogy háromujjú nyomai a kétlábú Theropoda dinoszauruszoknak (húsevők), kétlábú növényevőknek és madaraknak is lehetnek.75 Ez legalább megduplázza a háromlábú nyomokból szerzett paleontológiai információk volumenét anélkül, hogy túlságosan precízzé vagy kirívóvá tenné azokat.
Rejtélyes, hogy a legkorábbi dinoszaurusz-nyomokat a korai Triász üledékeiben (úgy 200 millió évesek)76 találták meg, miközben a legkorábbi dinoszaurusz csontokat a közép-Triász érára teszik, mintegy 20 millió évvel későbbre. Ez természetesen eléggé nyugtalanító a gradualista evolúció uralkodó dogmájára nézve, mert a dinoszauruszok eredetét a Perm idejére teszi, mintha túlélték volna az evolúció-történet legnagyobb fajkihalását, ami hozzávetőleg a Perm-Triász korok határán következett be. Emiatt egyes paleontológusok a nyomok kormeghatározásának elvetése mellett döntöttek (egyidejűnek tekintve őket az első dinoszaurusz-maradványokkal), míg mások kibővítették leszármazásukat, bevezetve egy új biológiai rendszertani egységet, az Archoszauruszokat, amelyekből állítólag a dinoszauruszok kifejlődtek. Az Archoszauruszokról úgy hiszik, hogy a dinoszauruszoktól láthatólag megkülönböztethetetlen nyomokat hagytak maguk után és azzal a kellemes jellemzővel bírnak, hogy nem hagytak fosszilis maradványokat valamint képesek magukba építeni a vitatott fosszilis formák előre- vagy visszafejlődését.
Egyes esetekben dinoszaurusz nyomokat találtak különböző egymást követő rétegekben, amelyre sokan a Teremtés könyve által leírt Özönvíz erős cáfolataként tekintenek. Ez természetesen attól függ, hogy alapként az áradás milyen forgatókönyvét vázoljuk fel. A gyorsan feltörő vizek globális áradásának lesznek heves, és ugyanígy nyugodt epizódjai is. Ésszerű feltételezni, hogy miután egész területek voltak befedve frissen lerakódott, kötetlen üledékekkel, a tengerszint viszonylagos változásai okozták az üledékek felszínre kerülését, elősegítve egyes dinoszauruszokat a partot érésben.77 A termékeny kreacionista író, Michael Oard megalkotta a BEDS kifejezést (Briefly Exposed Diluvial Sediments = Hirtelen előbukkant vízözönkori üledékek), hogy magyarázza vele a jelenséget.78 Ezeknek az átmeneti üledékes kőzetrétegeknek a felbukkanása vagy az ismételt lábnyomokat vagy az állatok hirtelen betemetődését vonhatta maga után vastag, finomállagú, vízmosta üledékekben. Később ennek eredményeként jelenhettek meg a felszínen a nyomokat tartalmazó üledékek. Ezért a legvalószínűbb, hogy világszerte közel az összes lábnyom alá volt vetve a vizes üledékek általi gyors betemetődésnek, összhangban egy világméretű Özönvízzel.
Az ember majomszerű elődei
A darwini evolúció összes nagyobb leszármazási vonalából az ember evolúciója van hozzánk időben a legközelebb és emiatt kövületek tömegének kellene ezt bizonyítania. De ez nem így van. Valójában jóval több dinoszaurusz fosszíliát van lehetőségünk tanulmányozni, mint feltételezett emberi ősöket. Ráadásul a fosszilis leletek többsége fogak és nem koponya formájában, , és még csak nem is különálló csontokként kerülnek elő.
Az elmúlt 50 év során a Homo sapiens evolúciós története drasztikusan megváltozott, a klasszikus fa-diagramból egy kusza bokorrá, majd egy gyümölcsöskertté alakult. Azonban ezek a látványos ábrák töredékességüket leszámítva sem láttatják az ember világos leszármazását (vagy az evolucionisták számára megfelelő kifejezéssel „felemelkedését”). Minden ilyen ábrázolás hemzseg a bizonytalanságoktól és a sok alternatív evolúciós nyomvonaltól, miközben az ábrák radikális változáson mentek keresztül az elmúlt évtizedekben. Az a benyomásunk támadt, hogy a sokféle „útvonalból” az egyéni preferencia alapján bármelyik kiválasztható, mivel Darwin nyomán mindegyik azt hivatott „bizonyítani”, hogy az ember „majomszerű ősökből” fejlődött tovább.79 Ahogy a Deinonychus esetében is különböző magyarázatok vannak arra, hogy „dinoszaurusz”-e vagy „madár”, az ember állítólagos evolúciójára is számos kövületet hoznak fel, amelyek tetszés szerint módosíthatók annak érdekében, hogy helyet készítsen az új felfedezéseknek. És a legtöbb új felfedezés valójában régi mintákból származik, melyeket évtizedekig tároltak fiókokban, hogy új kutatókkal újra-felfedeztessék egy későbbi időpontban. Az antropológia, jobban mint a tudomány más területei, reflektorfényért ácsingózik, amely pedig serkenti a finanszírozást.
Majomember a múltban
A Zinjanthropus boisei, más néven Diótörő Ember, egykor az ember majmokból való kifejlődésének népszerű „bizonyítéka” volt. Őt a híres paleo-antropológus, Mary Leakey fedezte fel 1959-ben, és egy jó évtizeden keresztül az evolucionista intézményrendszer erősen támogatta, mielőtt partvonalon kívülre került volna. Paranthropus-ra nevezték át, és ma már egy távolabbi rokonként, valamint az Australopithecina majmok egy tagjaként tekintenek rá, ami a majmok egy olyan kihalt, robusztus csoportja, melyek közé tartozott a híres Lucy is.80
A Ramapithecus az ember majomszerű rokonainak egy másik híres és hatásos példája, amelyet szintén kivontak a forgalomból. Őt a Nepálban az 1930-as évek elején felfedezett koponyatöredékek és fogak alapján győzedelmesen az emberi evolúciónak egy „pozitív bizonyítékaként” mutatták be. Azonban a későbbi, 1970-es években felfedezett újabb leletek alapján a Ramapithecus-t Silvapithecus-ra keresztelték át, és ma már csak egy kihalt majomfajként tartják számon, amely valószínűleg az orangutánok őse.
A Zinjanthropus-on és a Ramapithecus-on kívül legalább egy tucatnyi majomszerű fajt sorolhatnánk fel, amelyet valaha az emberi leszármazási vonal részeként kiáltottak ki, de mára már a szemeteskosárba száműztek. Akkor hol is van a bizonyíték?
A jelenkori majomember
Az Australopithecina fajokat, ideértve az A. afarensis-t, az A. africanus-t és az A. sediba-t, az ember és a majom közötti erős összekötő kapocsként tartják számon. Azonban a kiváló evolucionista anatómus, Charles Oxnard a fizikai antropológia Darwin-díjának címzettje nem fogadja el őket az ember őseiként. Amikor anatómiai jellemzőik az emberével és a ma élő majmokéval kerül összevetésre, akkor mindkét csoporttól messze távolabb esnek, mint amennyire azok egymástól.81
Az evolúciós leszármazás másik fontos ikonja a Homo habilis. Habár ember-ősként hirdették, ez azért ennél jóval nehezebb kérdés. Sokan úgy hivatkoznak rá, mint egy olyan képzeletbeli kategóriára, amelybe a különféle fajokhoz tartozó különböző fosszília-töredékek egyvelegét helyezik. És mit gondoljunk a Homo erectus-szal és a sok más majom-emberrel kapcsolatosállításokról? A hely szűke nem teszi lehetővé ennek a kérdésnek a teljes kifejtését, de az olvasó számára ajánljuk a creation.com/anthropology látogatását az összes ide vonatkozó ősleletről szóló részletes cikkekért.
A Neandervölgyi
Amióta a Neandervölgyit 1829-ben Belgiumban, 1848-ban Franciaországban és 1856-ban Németországban (a típuspéldányt) felfedezték, a mai ember ősének állították be. Aztán azt követően, hogy 1998-ban a Portugál Lagar Velho-nál felfedezték egy hibrid 4 éves gyermek csontvázát,82 az anatómusok felvetették annak a lehetőségét, hogy a Homo sapiens és a Homo neanderthalensis ugyanaz a faj. A genetikusok többsége erősen ellenezte ezt a lehetőséget az emberi genom evolúciós fejlődéstörténetébe vetett hitük alapján. További felfedezések, mint például a romániai Peştera cu Oase (A csontok barlangja)83 sem változtatta meg a genetikusok álláspontját. Azonban 2006 óta a Neandervölgyi Genom Projekt (NGP) egyre több genetikai bizonyítékot szolgáltat arra, hogy a Neandervölgyiek és az emberek bizony szaporodtak egymás között.84 Ahogy ezek a felfedezések tovább bonyolították (enyhe kifejezéssel) az emberi leszármazás hőn áhított fokozatos átmenetét, cikkek egész áradata vonta kétségbe az NGP érvényességét, amely természetesen válaszokat és viszont-válaszokat generált. Az antropológiai közösség ma megosztott a különböző nézetek között aszerint, hogy hol helyezkednek el a Neandervölgyiek az emberszabásúak evolúciós leszármazásán belül. Ez nemcsak a csontmaradványok, hanem a DNS tanulmányozásán is alapul. Habár nincs genetikai információ, amely alátámasztaná a Neandervölgyieket megelőző állítólagos emberi leszármazást, mégis úgy tűnik, hogy közmegegyezés alapján a Neandervölgyiek bekerültek az ember családjába.85
Általános jellemzőként az ember eredetét bemutató összes tekintélyes ábra rétegtani szempontból láttatja az egyes felfedezéseket, legtöbbjük között nincs összefüggés. Ha bármilyen kapcsolatot feltételeznek, pontozott vonallal jelzik – ami a bizonytalanság nyílt beismerése. A történet persze teljesen máshogy szól a médiában, bár ahol az emberi családtörténetet ilyen részletességgel mesélik el, úgy tűnik, mintha egy TV- családregényt látnánk!
Majomemberek, amelyek soha nem léteztek
A darwini evolúció hatását az emberiség eredetével kapcsolatos elképzelésekre nem szabad lebecsülni, különösen, mivel a gyarmatosítás és mindenféle rasszizmus indoklására használták, ahogy ezt az evolucionisták maguk is beismerték.86 Talán ez a magyarázata, hogy a félrevezetések miért szennyezték be az antropológia arculatát a kezdetektől – a majomemberek, amelyek soha nem léteztek. Az első nagyobb álhír a piltdowni ember volt (Eoanthropus dawsoni), amelyet 1912-ben a Londoni Földtani Intézetben olyan pompával mutattak be, mintha azok egy mai emberi koponya darabjai lennének. Lelkes tudósok újabb töredékeket és egy koponyát találtak ugyanabban a piltdowni sóbányában, Darwin angliai lakóhelyéhez közel. A maradványokat az elit brit tudósok a „legkorábbi britnek” titulálták:
Piltdown alátámasztott több, a fehér európaiak körében elterjedt rasszista nézetet. Az 1930-as és 1940-es években a kőzetrétegekben talált pekingi ember felfedezését követően, amely korban nagyjából megegyezik a piltdowni kőzettel, a Piltdownra alapozott törzsfejlődési fák és a fehér felsőbbrendűség ősi voltának megerősítése megjelent a szakirodalomban … Ha a piltdowni ember, a legkorábbi angolként volt a fehér fajok őse, miközben az emberré válás többi fokozatai a Homo erectus-ra vezetnek, akkor a fehérek átlépték a civilizáció vívmányainak küszöbét.87
1923-ban a német anatómus Frantz Weidenreich rámutatott, hogy a koponyatöredékek egy embertől származtak, de az állkapocs egy orangutáné volt reszelt fogakkal! Ezt követően a tudományos közösségnek 30 évébe tellett, hogy elismerje Weidenreich igazát.88 Ez volt az akkoriban általánosan elfogadott tudományos álláspont, de később mégis hibásnak bizonyult.
KÉP: A nebraskai ember fogának rajza. Balról jobbra: hátulsó, külső, elülső és belső nézőpontból.*
Smith, G.E., Hesperopithecus: the apeman of the Western world (Hesperopithecus: a nyugati világ majomembere), The Illustrated London News, p. 944, 24 June 1922.

KÉP:
William Jennings Bryan.
A nebraskai fog téves beazonosításával próbálta Osborn Jennings tekintélyét támadni.
A másik elhíresült példa egy az USA-ban, Nebraska államban talált fog, melyet a Hesperopithecus haroldcookii-hoz, ismertebb nevén nebraskai emberhez tartozónak írtak le.89 Részben ez a felfedezés szolgáltatta a hátteret az 1925-ös Scopes (majom) perhez. Habár a tárgyaláson az evolúció mellett egyetlen tényleges bizonyítékot sem vezettek elő, a tudós, Henry Fairfield Osborn, aki erről 1922-ben számolt be, felhatalmazva érezte magát, hogy gúnyt űzzön a bibliai álláspont védelmezőjéből, William Jennings Bryan-ből:
Tréfásan felvetették, hogy az állatot Bryopithecus-nak kellene elnevezni, Nebraska állam eddigi legelőkelőbb főemlős szülöttje után [politikusként Bryan kerülete Nebraska volt]. Mindenképp különös, hogy ezt a felfedezést 6 héttel azután (1922. március 5.) jelentették be, mikor a szerző azt tanácsolta William Jennings Bryannek, hogy nézzen utána a Jób könyve ezen szakaszának: „Beszélj a Földhöz és tanítani fog téged,” és az is egy jelentős egybeesés, hogy az első föld, amely erről beszélt, az a Nyugat-Nebraskában található Közép-Pliocén időszakból való Snake Creek homokos földje.90
Két évvel a pert követően a tekintélyes Science folyóirat elismerte, hogy a fog (amelyből az egész emberi fajt „rekonstruálták”) valójában egy kihalt pekari-félétől (egy sertéstől) származik.91 És ismét, a mindenkori elfogadott tudomány gúnyt űzött a bibliai teremtésből, majd később hamisnak bizonyult. Az igaz, hogy a tudomány gyakran korrigálja önmagát, azonban ezek a téves példák sokat tettek azért, hogy előmozdítsák az evolúciót a közgondolkodásban.
1972 augusztusában a National Geographic megjelentetett egy nagy cikket egy kőkori törzs (a Tasaday) felfedezéséről a Fülöp-szigeteki Mindanao-sziget őserdőiben. A NG egy dokumentumfilmet is készített, amelyet világszerte vetítettek a mozik. 1986-ban egy svájci újságíró és antropológus, Oswald Iten felfedte, hogy a Tasaday törzs története valójában egy gondosan kidolgozott álhír. Ezt a kormányügynökség korábbi vezetője, Manuel Elizalde92 követte el a kulturális kisebbségek védelme érdekében, aki pénzt akart gyűjteni a valódi Tasaday-ek életminőségének javításáért, akik nem egy kőkori törzs voltak, és a pénzt végül elsikkasztotta. 14 évbe tellett, hogy leleplezzék a hazugságot. Ez egy kis példa, de időközben hány embert befolyásolt, hogy elfogadja az ember evolúciós történetét, mélyen megalapozott tudományos ténynek gondolva azt?
2005-ben egy hatalmas botrány vezetett Rainer Protsch von Zieten professzor kényszernyugdíjazásához, aki történeteket talált ki és plagizált azért, hogy mutasson egy összekötő kapcsot az ember és a Neandervölgyi között.93 Habár a „felfedezéseket” a tudományos közösség mintegy 30 éven keresztül elfogadta, amikor a botrány kitört, sok antropológus próbálta megőrizni a tekintély látszatát azt állítva, hogy ők valójában nem hittek Protsch kutatásának, annak ellenére, hogy hivatkoztak rá. Ismét, ez volt napjaink elfogadott tudománya (és az akadémiai körök szemenszedett hazugságainak egy még frissebb példája).
Ez csak néhány a rengeteg eset közül, amit idézni lehet. Elgondolkodtató, hogy ha az evolúció igaz, miért van szükség ennyi csalásra?94 A válasz nyilvánvaló; a kövületek nem támasztják alá az ember és majom közös őstől való evolúcióját. A bizonyítékok hiányossága az, ami hajtja a frusztrált antropológusokat, hogy minden lehetséges módot felkutassanak, amivel ki tudják egyenlíteni a kövületek hiányát. A félrevezetések önmagukban még nem tekinthetők az evolúció Achilles-sarkainak, azonban rámutatnak egy fontos problémára: minden adatot, amelyet történelmi szempontból az emberi evolúció alátámasztására használnak, azóta már elvetettek. Ez az elutasítás (szerencsére) magában foglalja a félrevezető anyagokat, valamint a 19. és 20. századi majom-ember példákat is (amint korábban említettük).
Nyilvánvaló, hogy a „mainstream” tudományként bemutatott bizonyítékoknak csak egy töredéke megtévesztő. Azonban a változó történetek azt mutatják, hogy a magyarázatok ingadoznak, tehát óvatosnak kell lennünk, amikor különböző tényeket az ember leszármazása evolúciós paradigmájának „bizonyítékaként” jelölnek meg. Jó lenne azt gondolni, hogy a modern geológusok, paleontológusok és paleo-antropológusok bölcsebbek, és jobban meggondolják bejelentéseiket. Azonban ezek az emberek nem feddhetetlenek az evolúcióval kapcsolatos propagandatevékenységüket tekintve, amit az is mutat, ahogy kiszolgálták a média hisztériáját az Idára (amely egy lemur, és egy újabb „élő kövület”)95 és Ardira (egy olyan fosszília, amely „bejelentése” előtt több mint egy évtizeddel ismert volt)96 vonatkozó bejelentéseiknél, nem beszélve a jó öreg Lucy-t (majom) körülvevő őrületről.
Miért nincsenek emberi tetemek a dinoszauruszokéval együtt betemetve?
Mindig is vitathatatlannak tartották, hogy a fosszilis leletek világszerte azt az egyenletesen hierarchikus rendet követik, amely az egyszerűtől az összetett felé halad. Ennek a látásmódnak a része, hogy az ember a dinoszauruszok eltűnése után fejlődött ki, mintegy 60 millió évvel ezelőtt. Eszerint ez a magyarázata annak, hogy a dinoszauruszok és az ember maradványai miért nem találhatók meg ugyanabban a geológiai képződményben.
Kevéssé vonható kétségbe, hogy egy következetes fosszilis rendnek ne lenne valóságos alapja, azonban a rendhagyó kövületek nem teszik lehetővé, hogy egy ilyen sormintát geológiai vagy biológiai alapigazsággá léptessünk elő. Nos, ennek így is kellene történnie, ha a logika és a józanész nem lenne átitatva ideológiával! Természetesen a fosszíliák rendjére való rátekintésnek van egy másik módja is, amely helyet hagy a rendhagyó kövületeknek: a betemetődés rendje egy globális vízikatasztrófa által.
Először is, mivel a gyors betemetődés a fosszilizációnak egy nyilvánvaló ténye, a tengeri élőlényeknek volt a legnagyobb esélyük egy globális árvíz során fosszilizálódni, különösen a tengerfenék lakóinak, még inkább a letelepedett élőlényeknek. Ahogy korábban említettük, a fosszilis leletanyag nagy százalékát ezen állatok héjazatai teszik ki. A mai tengeri élőhelyek tekintetében a legtöbb tengerfenéken élő metazoa (többsejtű állat) egyszerű organizmus, miközben a komplexebb élőlények többsége a felszínhez közel szeret élni. Természetesen mindkét esetre vannak kivételek (egyszerűbb élőlények a felszínen úsznak [zooplanktonok] és emlősök [óriás ámbráscetek] nagyon mélyen úsznak), a biota valódi sűrűségét tekintve a tengerfenéken lakók túlnyomórészt egyszerűbb élőlények (kagylók, korallok, tengeri csillagok, zsákállatok stb.).
Másodszor, a felszínen élők egy része, különösen a tengeri emlősök, túlélhették az Özönvizet, és azok, amelyek elpusztultak, kevesebb esélyük lehetett fosszilizálódni, mivel szétbomolhattak anélkül, hogy betemetődtek volna, feltéve, ha túlélték a nagy üledékképző időszakokat. Így többnyire a fosszilis leletréteg felsőbb részeiben lennének jelen, azonban további kivételek (lásd a fentebb bemutatott rendhagyó kövületeket) várhatók.
Harmadszor, az Özönvíz előtti emberi populáció-geográfiai eloszlása és mérete nagy szerepet játszhatott annak meghatározásában, hogy hol, mikor és mennyi ember temetődjön be fosszíliaként. Azonban még mindig keveset tudunk az Özönvíz előtti föld domborzatáról, nem tudjuk hol éltek az emberek (tengerparton vagy messze a tengertől?), és nem tudjuk, hogy hány ember élt, amikor eljött az Özönvíz. Úgy tűnik, hogy a Földön jelentős kiterjedésű, alacsony fekvésű, mocsaras területek voltak, amelyre sok olyan növény és állat szolgál bizonyítékként, amelyekről úgy tudjuk, hogy ilyen jellegű környezetben éltek. Ebben az esetben az emberek magasabb helyeken élhettek, messze a tengertől, és messze a növényevő dinoszauruszoktól, amelyek nem illettek volna bele az ember mezőgazdasági tevékenységébe. Azonban ez olyasvalami, amit egyszerűen nem tudhatunk bizonyosan.
Az emberi népességről, akik az Özönvíz eljövetelekor éltek a Földön, szintén nincs ismeretünk. A különböző becslések a néhány százezertől egészen a milliárdig terjednek. Lévén, hogy emberi leletek ritkák vagy teljesen hiányoznak az üledékrétegekben, az alacsonyabb becslések valószínűbbnek tűnnek. Ádámtól Noéig 10 férfigeneráció és több mint 1500 év telt el. Ez tág teret enged egy nagy emberpopulációnak, azonban a Biblia a világot erőszakkal telinek írja le (1. Mózes 6.), tehát lehetséges, hogy a népesség nem volt olyan nagy, mint amilyen lehetett volna a gonoszság nélkül.
Negyedszer, a szárazföldi emlősök magasabban fekvő helyekre szökhettek (attól függően, hogy milyen gyorsan érte el az árvíz a földet), sokan közülük (a kagylókkal ellentétben) változó hőmérsékletű helyekre tudnak úszni. Ily módon feltételezhető róluk, hogy az utolsó olyan élőlények között voltak, amelyeket teljesen beborított a víz, és az árvíz emelkedésekor szintén utoljára temetődhettek be. Miután az árvíz szintje elérte a csúcsot, a víz elkezdett lefolyni a szárazföldekről a tengerekbe, elkoptatva a kontinensek akkori felszínét. Az eróziónak ezt az időszakát a geológusok helyesen ismerték fel, akik egyes helyeken ezt a folyamatot „a nagy tarolásnak” nevezték el. Helyenként mérföldeket beborító üledék mosódott el. Az élőlények maradványainak jó része később az Özönvíz során temetődött el, és ebben az időben az üledékekkel együtt mállottak szét.
Ötödször, a kérdés könnyen megfordítható és így elég kényelmetlenné válik az evolucionisták számára. Például tudjuk, hogy a bálnák és a maradványhalak ma is az óceánokban élnek, de nem találtak bálnákat a maradványhalak kövületeivel együtt eltemetve. Valójában nincsenek maradványhal kövületek az állítólagos 65 millió évesnél fiatalabb üledékekben, amikor a fosszilis leletanyagból eltűntek, egészen mostanáig. Miért nem látjuk őket betemetve azokkal a tengeri lényekkel együtt, amelyek valamikor ezen időszak alatt éltek? Az evolucionista közösség válasza erre így szól: „A fosszilis leletanyag hiányos.” De akkor miért nem mondhatják a bibliai kreacionisták, hogy „Az Özönvíz nem őrizte meg tökéletesen az Özönvíz előtti világot?”
Erős a meggyőződésem, hogy ha bármikor egy emberi koponya kerülne elő, mondjuk egy dinoszaurusz-maradványt tartalmazó rétegből, az evolucionisták valamilyen előre legyártott vagy nehezen megfogható magyarázattal rukkolnának elő egy ilyen problémás leletre.
Következtetések
E fejezet során gondolatban egynél több paradigma segítségével tekintettem a fosszilis leletanyagokra. Amíg az evolúciós feltételezéseken alapuló magyarázatok számtalan kifejezési módra, kutatási munkaórára és jelentős anyagi támogatásra leltek, addig a kreacionista, Özönvíz utáni korszakra vonatkozó magyarázatok több mint két évszázadig perifériára voltak szorítva. Ezen felül, az elmúlt fél évszázadban szüntelen, jól megszervezett, agresszív támadások célpontjaivá váltak. Az evolucionista média abszolút dominanciájának köszönhetően az általános közvélemény számára csak elvétve mutattak be a Teremtés mellett szóló érveket, és csak azzal keltettek feltűnést, hogy a kreacionisták még léteznek. Ha mégis meg lettek említve érvek, akkor ezek torzként, idejétmúltként vagy logikai tévedésként lettek bemutatva, “szalmabáb” érvelésnek nevezve ezeket. Azon kevés alkalommal, amikor egy fiatal földben és az Özönvízben hívő geológusnak engedélyeztek egy kis megjelenést a médiában, gyakran kötelességüknek érezték, hogy mellétegyenek egy evolucionista geológust, aki képviselte az ellenoldalt. Azonban az ellenfél gyakran nem fogadta el, hogy egy fiatalföld-hívő geológust kértek meg arra, hogy a médiában mutassa be nézeteit az evolucionista geológussal szemben.
A Bibliai alapokon álló geológusoknak még mindig sokat kell kutatniuk a fosszilis leletanyagban, hogy megértsék a világméretű Özönvíz hatásait és folyamatát. Ez még nincs teljesen kidolgozva, ami annyiból jó, hogy bőséges tennivalót hagy a jövő generációi számára. Ugyanakkor az is tény, hogy bár a bibliai geológusoknak fizikai hozzáférésük van a fontos geológiai helyszínekhez, azokhoz is, melyeket “aranyrögként” aposztrofálnak, mégis arra kényszerülnek, hogy az evolucionista geológusok által gyűjtött adatokat értékeljék át, amelyek evolucionista értelmezésekkel vannak megterhelve. Miután Francis Bacon a Novum Organum c. művében (1620) azt vetette fel, hogy az adatokat részrehajlás nélkül, induktív módon kell gyűjteni, ill. magyarázni, világossá vált, hogy az ilyen megközelítés nem reális, mivel az „adatgyűjtők” ezt a titulust részrehajlásuk halmozásával szerezték, és ezt még oktatásnak is nevezték! A geológiai helyszínek újra-meglátogatása általi átértelmezések szükségesek, de ez egy lassúbb folyamat.
Átvitt értelemben azt is mondhatjuk, hogy a kövületek azok a betűk, melyeket a geológusok az őket tartalmazó sziklák megfejtésére használnak. A legtöbb mai geológus elutasítana egy ilyen állítást, mégis az ő „bibliájuk”, amely vonakodva ugyan, de beismeri:
… a mai napig hittétel a földtan tudósai számára, hogy a fejlődő nemzetközi rétegtani skála beosztásait a kőzetek fosszilis tartalma határozza meg. Ennek végig vitele azonban nehézségekhez vezet: a határok az új felfedezésekkel változhatnak; a részleges fosszíliák alapján meghatározott határok történeti jellegűvé válhatnak; nézeteltéréshez fog vezetni, hogy melyik rendszertani kategória legyen a meghatározó.97
Más szavakkal, a kőzeteket továbbra is fosszilis tartalmuk alapján sorolják be, még akkor is, ha ez nehézségekhez vezet. Az evolucionista geológus számára a nehézségek abból adódnak, hogy a kövületeket nem önmagukban értelmezik, hiszen ez egyszerűen olyan kockázat volna, amelyet nem éri meg felvállalni, mivel tágra nyitja a zsilipeket (szó szerint!). Ez adja a lehetőségetarra, hogy sok kőzetképződményt rövid időkeretekbe lehet besűríteni a rétegtan egyszerű alkalmazásával. Valójában a legutóbbi évtized egy nagy váltást hozott az üledékképződés magyarázatában:
… tisztában kell lennünk az üledékképződésben tátongó hosszú távú idő-hézagokkal, valamint azzal a ténnyel, hogy a legtöbb környezetben mind a nem-lerakódó, mind az erodálódó területek szélesebb körben elterjedtek és állandóbbak, mint a tisztán lerakódó területek … Még a mély tengerekben is, melyekre egykor a nyílttengeri üledékekre való szakadatlan hóesésként gondoltunk, ma már hatalmas töréseket fedezünk fel a lerakódásban, sőt még az erózióban is.98
Másként fogalmazva, ahogy a kőzetek mozognak, a geológiai idő nagyobbik része a rétegek közötti résekbe préselődik, és emiatt a kövületek számítanak a legüdvözöltebb „időhorgonynak” (megnyugvást adva az evolucionistáknak). Kövületek nélkül a geológiai oszlop a kőzetek egymás után következő száraz, érdektelen lajstroma lenne, hébe-hóba kiegészülve újabb kőzetekkel.
Rövid értekezésünkben szóltunk a rendhagyó kövületekről, az átmeneti formákra vonatkozó komoly hiányosságokról, és az evolucionista tudomány kudarcáról, hogy nem képes az embernek olyan majomszerű őspéldáit felmutatni, amelyek kiállnák az idő próbáját. Ebből láthatjuk, hogy a fosszilis leletanyag az evolúciótannak ma is ugyanúgy Achillesz sarka, mint volt Darwin idejében.
Hova vezet ez?
Az első két fejezet együtt a darwinizmus „szent Gráljával”, a természetes szelekcióval és kiegészítőjével, a genetikával foglalkozik. A harmadik fejezet az élet eredetére vonatkozó modellek kémiai és statisztikai megkötéseiről szól, amely kéz a kézben jár a kövületekkel. Nemcsak az életnek az élettelen anyagból való származásának koncepciója nehézkes, és nemcsak kémiailag és fizikailag is lehetetlen, hanem még a hosszú időtávú diverzifikációt sem tükrözi a fosszilis leletanyag!
Még az élet kezdetének állítólagos kifejlődése után sincs nyoma abban a rengeteg kísérletben annak, hogy az életnek szükségszerűen a mai állapotra kellett eljutnia. Továbbá a természetes szelekció és a genetika első fejezetei alapján sem létezik olyan mechanizmus a fizikai világban, mely az életnek olyan mérhetetlen változatosságához és komplexitásához vezethetett volna, mint amit ma látunk. Az első két fejezet azzal foglalkozik, amit Darwin igaznak gondolt. A következő kettő azt tárgyalja, amiről feltételezte, hogy igaz, és amiről azt gondolta, hogy a jövőbeli felfedezések igazolni fogják. Innen már látjuk, hogy Darwinnak mire volt szüksége, hogy igaz legyen: a különösen hosszú időszakok. A fosszilis leletektől a kőzetanyag kitárgyalása felé mozdulunk el, függetlenül attól, hogy ez megadja-e a Darwinnak szükséges időt, avagy sem.
__________________________________
Lábjegyzet:
1. Becenevét az Anglia nagy részén található kőzetrétegek kiterjedésével és helyszíneinek átfogó térképezésével összefüggésben kapta.
2. C. Darwin: A fajok eredetéről, 1. kiadás, 6. fejezet, 171.o., John Murray, London, 1859.
3. A „Golden spike” („aranyrúd”) a Global Boundary Stratotype Section and Point-nak (GSSP) egy köznyelvi változata, amely a Nemzetközi Stratigráfiai Bizottság által elfogadott fizikai jelzőeszköz a geológiai oszlop részei közötti határok megállapítására.
4. A Phanerozoikum, görög jelentése „látható élet”, az evolúciós időskála szerinti „korszakokat” fedi le az élet bőséges fosszilis bizonyítékaival.
5. S.J. Gould: Wonderful Life (Csodálatos Élet), W.W. Norton & Co., New York, 56.o. 1990.
6. D. Wavey és tsai: A 3,4 milliárd éves közetben talált kén-metabolizáló sejtek mikrofosszíliái Nyugat-Ausztráliában, Nature Geoscience 4:698-702, 2011.
7. W.J. ReMine: Desperate attempts to discover „the elusive process of evolution” (Kétségbeesett próbálkozások „az evolúció kétséges folyamatának” felfedezésére): A review of The Altenberg 16: An Exposé of the Evolution Industry by Suzan Mazur, J. Creation 26(1):24-30, 2012; creation.com/review-altenberg-16.
8. M. Syvanen: Evolutionary implications of horizontal gene transfer (A horizontális génátvitel evolúciós összefüggései), Annual Review og Genetics 46:339-356, 2012.
9. P. Jerlström: Is the evolutionary tree turning into a creationist orchard? (Az evolúciós fa kreacionista gyümölcsöskertté válik?) J. Creation 14(2):11-13, 2000; creation.com/creationist-orchard.
10. S. Doyle: Kladisztika, evolúció és a fosszíliák, J. Creation 25(2):32-39, 2011; creation.com/cladistics.
11. Más források 3,5 milliárd évre helyezik; 100 millió év elvétele vagy hozzáadása megszokott dolog ilyen nagyságrendben.
12. C. Wallis: Az evolúciós háborúk, 2005. augusztus 7.; time.com
13. T. Walker – Slow Fish in China, Creation 22(3):38-39, 2000; creation.com/slow-fish-in-china.
14. S.J. Sameeni: The Salt Range: Pakistan”s unique field museum of geology and paleontology (A Sóbarlang: Pakisztán egyedülálló geológiai és paleontológiai múzeuma, in Lipps, J.H. and Garnier, B.R.C. (kiadások), PaleoParks-fosszilis lelőhelyek védelme és megőrzése a világ minden táján, Carnets de Géologie/Notebooks on Geology, Brest, Book 2009/03, 2009.
15. R.V.V. Anderson: Tertiary stratigraphy and orogeny of the northern Punjab (Észak-Punjab harmadkori stratigráfiája és hegyképződése), Bull. Geol. Soc. America 38:665–720, 1927.
16. A Harmadkor az evolúciós geológiai oszlop harmadik leghosszabb szakasza, a „dinoszauruszok kora” után.
17. B.Sahni: Age of Saline Series in the Salt Range of the Punjab (Sólerakódások kora a Punjabi Sóbarlangban), Nature 153:462–463, 1944.
18. J. Coates és tsai: Age of Saline Series in the Punjab Salt Range (Sólerakódások kora a Punjab-i Salt Range-ben), Nature 155:266–277, 1945.
19. B. Sahni: Microfossils and the Salt Range Thrust (Mikrofosszíliák és a sóhegyvonulat-gyűrődés), Proceedings of the National Academy of Sciences, India, Section B, (Az Indiai Nemzeti Tudományos Akadémia Kiadványai, B cikk) vol. 16:i–xlx, 1946.
20. Sahni, Microfossils and the Salt Range Thrust, pp. xlv–xlvi.
21. Yeates, R.S. et al., Late Quaternary deformation of the Salt Range of Pakistan (A Pakisztáni Sóképződmény késő negyedkori torzulása), Geol. Soc. of Am. Bull. 95:958–966, 1984; Jaumé, S.C. and Lillie, R.J., Mechanics of the Salt Range-Potwar Plateau, Pakistan, Tectonics 7:57–71, 1988.
22. Butler, R.W.H. et al., Salt control on thrust geometry, structural style and gravitational collapse along the Himalayan Mountain Front in the Salt Range of Northern Pakistan (A Himalájai hegységfront sószabályzása a lemezmozgás geometriájában, szerkezeti modellje és gravitációs összeomlása az észak-pakisztáni sóképződményben) ; in Lerche, I. and O”Brian, J.J. (eds), Dynamical Geology of Salt and Related Structures (A só és a hozzá kapcsolódó struktúrák dinamikus geológiája), Orlando, Academic Press, pp. 339–418, 1987.
23. Sameeni, The Salt Range.
24. R.M. Stainforth: Occurrence of pollen and spores in the Roraima Formation of Venezuela and British Guiana (Pollenek és spórák megjelenése a Venezuelai és a Brit-Guyana-i Roraima Képződményben), Nature 210:292–294, 1966.
25. E. Silvestru és C. Wieland: A pollen-paradoxon, 2011.június 27.; creation.com/pollen-paradox.
26. Az eddig felfedezett legidősebb gerinces fosszília, Discovery News, 2003.október 23.; dsc.discovery.com.
27. Egy farmer fosszília felfedezése kelt izgalmat, BBC News, 2003. október 24., news.bbc.co.uk.
28. S. Leclerq: Vaszkuláris növényekre vonatkozó bizonyítékok a Kambriumban, International Journal of Organic Evolution 10(2):109–114, 1956.
29. K. Jacob és trsai: A Kambriumban élt növények létezésére vonatkozó bizonyítékok, Current Science 22:34–36, 1953.
30. P.F. Hoffmann és társai: A Neoproterozoic snowball Earth, Science 281:1342–1346, 1998.
31. Horner, J.R., Upper Cretaceous dinosaurs from the Bearpaw Shale (marine) of South Central Montana, with a checklist of Upper Cretaceous dinosaur remains from marine sediments in North America, (A Dél-Közép Montana-i (tengeri) „Medvemancs Palából” származó Felső-Krétai dinoszauruszok, kiegészítve az Észak-Amerikai tengeri üledékeiben található Felső-Krétai dinoszauruszok ellenőrzőlistájával) J. Paleontology 53(3):566–577, 1979.
32. Bakker, R.T., The Dinosaur Heresies (Dinoszaurusz eretnekségek), Zebra Books, New York, p. 40, 1986.
33. Everhard, M.J. and Hamm, S.A., A new nodosaur specimen (Dinosauria: Nodosauridae) from the Smoky Hill Chalk (Upper Cretaceous) of western Kansas (Egy új Nodoszaurusz faj, amely a Nyugat-Kansas-ben található Smokey Hill (felső-krétai) mészkőjéből származik) , Transactions of the Kansas Acadamy of Science 108:15–21, 2005.
34. Drumheller Mail, April 1st, 2011.
35. Horner, Upper Cretaceous dinosaurs from the Bearpaw Shale. (Felső-Krétai dinoszauruszok a „Medvemancs Palából”.
36. Darwin: Fajok eredete, 9. kiadás
37. Gould: Csodálatos Élet 57. o.
38. Gould: Csodálatos Élet 210–211. o.
39. Gould: Wonderful Life (Csodálatos élet), 46.o.
40. Gould, Wonderful Life (Csodálatos élet), 230.o.
41. Gould, Wonderful Life (Csodálatos élet), 230-231.o.
42. M. Schweitzer és társai: Lágyszövetek és sejtszintű megőrződés a Tyrannosaurus rex-ben, Science 307:1952–1955, 2005.
43. M.H. Armitage és K.L. Anderson: A Triceratops horridus dinoszaurusz szarvából hátramaradt fosszíliának lágyszövetes csontja, Acta Histochemica 115(6):603–608, 2013.
44. G. Kaye és társai: A dinoszaurusz lágyszövetek bakteriális biofilmként vannak interpretálva, PLoS ONE 3(7):e2808.
45. D. Vergano: A tegnap T.Rex-e a ma csirkéje, USA Today, 2007.április 12.; usatoday.com.
46 J.C.Pearce: A Wiltshire-i Christian Malford közelében, a Nagy Nyugati Vasút építése során, az Oxford Clay rétegeiben felfedezett ammoniták szájáról és fosszíliáikról, Proc. Geol. Soc. London 3:592–594, 1841.
47. A tafonómia a fosszilizációt magába foglaló folyamatok tanulmányozása a haláltól a fosszíliák helyreállításáig.
48. P. R. Wilby és társai: Az Oxford Clay Formation-ban (Jurassic, England) megtalálható puhatestű lábasfejűek felhalmozódásának tafonómiája és eredete, Paleontology 47(5):1159–1180, 2004.
49. Tinták találtak egy Jurassic korszakból származó tintahalban, BBC News, 200ö.aug.19.; news.bbc.co.uk.
50. J. L. Franzen és társai: A német Messel városában talált Közép-eocén korabeli teljes főemlős csontváz: morfológia és paleobiológia, PLoS ONE 45(5):e5723, 2009.
51. D. Batten: A túlreklámozott darwini fosszíliák, 2009.május 23., creation.com/ida.
52. Henneberg, M., The Hobbit Trap, Wakefield Press, Kent Town, Australia, p. 25, 2008.
53. X. Xu és társai: Egy Kínában talált Archaeopteryx-szerű therapoda és az Avialae eredete, Nature 475:465–470, 2011.
54. Tollas dinoszauruszok, A Blanding-i Dinoszaurusz Múzeum, Utah állam, USA – A Royal Ontario Múzeum kiállítása, 2005.
55. T. Lingham-Soliar: A toll evolúciója: Sinosauropteryx, egy állítólag tollas dinoszaurusz élete, halála és megkövülése, J. Ornithology 153(3):699–711, 2012. Ez a szerző olyan előzetes állításokat is tesz, hogy a feltételezett melanoszómák (amelyek a tollakban lévő színeződés jelzői lehetnek), amelyeket egyes dinoszaurusz-kövületekben fedeztek fel, olyan leletnek a nagyítása volt, mely már eleve a használt mikroszkóp részét képezte. Lásd még: J. Sarfati: ‘Tollas’ dínók: mégsincsenek tollak! 2012.Július 24.; creation.com/featherless.
56. E.B. Daeschler és társai: Egy devoni tetrapoda-szerű hal és a tetrapoda testfelépítés evolúciója, Nature 440:757–763, 2006., Lásd még: J.Sarfati, Tiktaalik roseae—egy halszerű ‘missing link’, 2006; creation.com/ tiktaalik.
57. G. Niedzwiedzki és társai: Tetrapoda ösvények a korai közép-devoni időszakból Lengyelországban, Nature 463:43–48, 2010., Lásd még: T.Walker, A híres hal-lelet valóban teljesen ép?; creation.com/tiktaalik-finished.
58. N. Eldredge: Darwin újrafelfedezése, Orion Publishing, London, p.95, 1996.
59. N. Rybczynski, M.R.Dawson és R.H.Tedford: A félig tengeri sarkvidéki emlős húsevők a Miocén korból és a Pinnipediák eredete, Nature 458:1021–1024, 2009.
60. R. Dawkins: A Föld legnagyobb mutatványa, Bantam Books, New York, 2009, 173.o. Dr Jonathan Sarfati más olyan fosszíliákkal is foglalkozott, amelyeket Dr.Dawkins átmeneti formáknak titulált a The Greatest Hoax on Earth (A Föld legnagyobb félrevezetés-ében, Creation Book Publishers, 2010 (elérhető a creation.com-on). Lásd még: creation.com/fossils.
61. D. Batten: Egy újabb fontos összekötő láncszem mond csődöt: Puijila, az úszólábúak állítólagos őse egy vidra, Creation 35(1):51–53; creation.com/puijila. Részletesebb indoklásért lásd az alábbi cikk kiegészítését:
C. Werner: A Puijila “családfájának” elemzése, in Nature 2009, creation.com/puijila-s.
62. C. Werner: Evolúció: A nagyszabású kísérlet, 1.kötet, New Leaf Press, Green Forest, Arkansas, 225.o., 2007 (a creation.com-on keresztül elérhető).
63. D. Batten: Hamis bálnaevolúció: egy újabb evolúciós szimbólum semmisült meg; creation.com/whale-evolution-fraud, 2014.április 12.
64. A. Williams és J.Sarfati: Egyáltalán nem olyan, mint egy bálna, Creation 27(2):20–22, 2005; creation.com/not-at-all-like-a-whale.
65. D. Batten: Rodhocetus és a bálnaevolúció egyéb történetei, Creation 33(3):54–55, 2011; creation. com/rodhocetus.
66. N. Eldredge: Time Frames: The Rethinking of Darwinian Evolution and the Theory of Punctuated Equilibria, (Időkeretek: A Darwini Evolúciónak és a Punctuated Equilibrium elméletének újragondolása), Simon & Schuster, New York, pp. 188–189, 1985.
67. A legidősebb talált tarisznyarák fosszília 445 millió éves, ScienceDaily, 2008.február 8.; sciencedaily. com.
68. D. Wallosek és társai: A Kínai Hubei tartományban talált skorpió a Felső-Devoni időszakból (Arachnida, Scorpionida), N. Jub. Geol. Paläont. Mh. H3:169–180, 1990.
69. A. J. Jeram: A Sziluri és a Devoni skorpiók fejlődéstörténete, kategorizálása és evolúciója, Kiadványok a 17. századi régészeti értekezésekből, Edinburgh, 1997; european-arachnology.org.
70. I. Anderson: Fenyő ‘dinoszaurusz’ lurks rejtőzik egy hasadékban, New Scientist 1957:5, 1994.
71. D. Catchpoole: ‘A dinó-fa’ projekt befejeződik, 2009.júl.2.; creation.com/dino-tree-project-ends.
72. P. Cartwright és társai: Kivételes módon megőrződött medúzák a Közép-Kambriumi időszakból, PLoS One 2(10):e1121, 2007. Lásd még: D. Catchpoole: Kivételes módon megőrződött medúzák, Creation 30(4):21, 2008; creation.com/epjelly.
73. M.G. Lockley és társai: Dinoszaurusz tó: A Purgatoire völgybeli Tracksite Area-n található Dinoszaurusz története, Colorado Geológiai Tanulmány Rendkívüli Kiadás 40, Denver, CO, USA, 35.o., 1997.
74. A. Hunt és társai: Az Új-Mexicói (Felső-Pennsylvaniai) Cutler Group szikláiban talált hatalmas Arthropleura nyoma, a Diplichnites Cuithensis, Az Amerikai Geológiai Közösség 2004-es Denver-i Éves Találkozója; gsa.confex.com.
75. M.G.Lockley és C.Meyer: Európa Dinoszaurusz nyomai és más fosszilis lábnyomok, Columbia University Press, New York, 2000.
76. M.G. Lockley és társai: Dinoszaurusz-tó: A Purgatoire völgybeli Tracksite Area-n található Dinoszaurusz története, Colorado Geológiai Tanulmány Rendkívüli Kiadás 40, Denver, CO, USA, 67–69.o., 1997.
77. M.J.Oard: Dinoszaurusz kihívások és rejtélyek, Creation Book Publishers, Atlanta, GA, USA, 113– 128.o., 2011 (a creation.com-on keresztül elérhető).
78. M.J.Oard: A K/T az Özönvíz utáni határvonal?—2.rész: ősklímák és fosszíliák, J. Creation 24(3):87– 93, 2010; creation.com/kt-boundary-flood-2.
79. C.Darwin: Az ember leszármazása és kiválogatódása a nemiséggel összefüggésben, 1.kiadás, John Murray, London, 86.o., 1871.
80. A mítoszgyártás nyomában: A képmás ereje, Creation 21(4):4, 1999; creation.com/myth-making. Lásd még: R.Grigg: Hiányzó nyom: Hogyan idézte elő egy misszionárius család a “majomember” kutatás egyik legfőbb arcának (Louis Leakey-nek) a felfutását? Creation 26(3):24–27, 2004; creation.com/leakey.
81. C.E.Oxnard: Az australopithecinek helye az emberi evolúcióban: indokolt kételyek? Nature 258:389-395, 1975.
82. C.Duarte és társai: A Portugáliai Abrigo do Lagar Velho-ból származó, korai Felső-Paleolitikumi emberi koponya és a mai ember kifejlődése az Ibériai-Félszigeten, Az USA Nemzeti Tudományos Akadémiájának Kiadványai 96(13):7604–7609, 1999.
83. E.Trinkaus és társai: A Romániai Pestera cu Oase-ből származó kora-modern ember, Az USA Nemzeti Tudományos Akadémiájának Kiadványai 100:11231-11236, 2003.
84. R.E.Green és társai: A Neandervölgyi genomjának szekvencia-terve, Science 328:710–722, 2010; K.Than: A Neandervölgyi és a mai ember egymással való szaporodása – az első stabil DNS-bizonyíték, National Geographic Napi Hírek, 2010. május 6.; news.nationalgeographic.com.
85. R.W.Carter: A Neandervölgyi genomja a mienkhez hasonló (Előfordulhatnak Neandervölgyiek a következő családi összejöveteleden!), 2010. június 1., creation.com/neandergenes.
86. Lásd a 8. fejezetet: Etika és moralitás.
87. S.J.Gould: A panda hüvelykujja, Norton Press, New York, 117.o., 1980.
88. A.J.White: A Piltdowni ember, 2003. november 24.; creation.com/the-piltdown-man-fraud.
89. A.Sibley: Egy gyors pillantás a Nebraska-i emberre, J. Creation 22(2):108–113, 2008; creation.com/nebraska2.
90. H.F.Osborn: A Föld Bryan-hez szól, The Forum 73:796–803, 1925.
91. W.K.Gregory: A Hesperopithecus látszólag se nem majom, se nem ember, Science 66:579–581, 1927.
92. O.Iten: A Tasadayok: Egy Fülöp-szigeteki kőkorszaki átverés, Neue Zürcher Zeitung, Zürich, 77–89.o., 1986. április 12.
93. Például lásd L.Harding: A modern ember történelme rávilágít, hogy a német tudós csalónak bizonyul, 2005.február 18.; guardian.co.uk.
94. J.Bergman: A csalás ragályossága miért van még ma is jelen a tudományban, J. Creation 18(3):104–109; creation. com/science-fraud-epidemic.
95. D.Batten: Ida: az agyonpropagált Darwin-fosszília, Creation 32(1):44–46; creation.com/ida.
96. C.Wieland: Megint az Ardipithecus: egy újrahasznosított majom-ember , 2009.október 5., creation.com/ardipithecus-again.
97. F.M.Gradstein és társai: A kronostratigráfia: az idő és a kövek összekötő eleme; Gradsteinben, F.M., Ogg, J.G. and Smith, A.G. (kiadások), A Földtani Időskála 2004, Cambridge University Press, 20.o., 2004.
98. H.G.Reading és B.K.Levell: Az üledékes kőzetleletek vizsgálatai; Readingben, H.G.(kiadés.), Üledékes környezetek: Folyamatok, képződmények és stratigráfia, Blackwell Science, 18.o., 2002.