EAH 1. Természetes szelekció
Részlet az Evolúció Akhilleusz sarkai (halálos pontjai) című könyvből

1. fejezet:
Természetes szelekció
15-47 oldal
A könyvet Robert Carter, Ph.d szerkesztette.
Kiadó: Creation Book Publishers. Második kiadás, 2014. október
Honlap: Creation.com
Fordította: Dr. Cserháti Mátyás biológus.
Lektorálta: Dr. Hecker Kristóf biológus.
Dr Don Batten Ph.D: Az evolúció Akhilleusz sarkai című könyvének fejezetenkénti fordítása.
Tartalomjegyzék:
1. fejezet: Természetes szelekció
6. fejezet: Radiometrikus kormeghatározás
8. fejezet: Etika és erkölcsiség
- fejezet
Dr. Donald Batten
Ph.D. Növényfiziológia, Sydney Egyetem
Dr. Donald Batten a trópusi növénytermesztés szakértője, különösen a mungóbab, licsi, guáva, cukoralma, illetve a mangó termesztéséhez ért. Nem ismeretlen a tudományos élet számára, hiszen Don írásai mind világi, mind keresztyén folyóiratokban rendszeresen jelennek meg, és több tudományos cikket és könyvet is jegyez. Behatóan foglalkozott a globális jelentőségű termesztett növények környezeti adaptációval, környezeti fiziológiájával és ásványi táplálkozásával, így kiválóan meg tudja ítélni a természetes szelekció jellemzőit, mivel munkája során állandóan ezzel foglalkozott. Több évtizednyi tapasztalatát, amit mint tudós és kreacionista szerzett, az „Evolúció Achilles-ina” című könyvében írja meg.
Lásd: creation.com/dr-don-batten
A természetes szelekció: a darwini evolúció sarokköve
Charles Darwin 1859-es könyvének teljes címe a természetes szelekció fogalmát fejezte ki: A fajok természetes kiválasztással való eredete, avagy a sikeres fajok fennmaradása a létért folyó küzdelemben. Vagyis a „természet” azokat az egyedeket őrizte meg, amelyek a legjobban alkalmazkodtak a környezetükhöz.
A természetes szelekció egy józan, ésszerű eszme. Azok az élőlények, amelyek olyan tulajdonságokkal rendelkeznek, amikkel egy adott környezetben jobban túlélnek, fennmaradnak azokkal szemben, amelyek nem rendelkeznek ezekkel a tulajdonságokkal. Például azok a farkasok, amelyek kis fülekkel, rövid lábakkal és vastag bundával rendelkeznek, jobban túlélnek a sarkvidéken, mint azok a farkasok, amelyeknek nagy a füle, hosszúak a lábai, és vékony a szőrzetük. Ezek a különbségek az állatok azon képességeit érintik, hogy tudnak-e hőt felvenni vagy leadni; ezek fontosak a hideg, illetve a meleg éghajlaton.
Noha a természet nem egy tudatos lény, és ezért nem tud „válogatni”, a természetes szelekció kényelmes kifejezés arra, hogy az egyedek és a gének időben és különböző környezetben való túléléséről beszéljünk. 1868-ban Darwin tisztázta, hogy a természetes szelekciónak nem volt iránya vagy célja:
Az életért folytatott küzdelemben azon változatok megőrzése, melyek akár szerkezetükben, összetételükben vagy ösztöneikben előnyt biztosítanak, az, amit természetes szelekciónak neveztem el. Ugyanezt az alapelvet nevezte Herbert Spencer „a legrátermettebb túlélésének”. A „természetes szelekció” bizonyos esetben szerencsétlen megfogalmazás, mivel tudatos választást sugalmaz; de ettől eltekintünk némi hozzáértés után … A rövidség kedvéért néha a természetes szelekcióról úgy beszélek, mint egy intelligens erőről – ugyanabban az értelemben, mint ahogy a csillagászok arról beszélnek, hogy a gravitáció a bolygók mozgására hat, vagy ahogyan az állattenyésztők arról beszélnek, hogy az állatokat szelekció révén háziasítani tudják. Sem az egyik esetben, sem a másikban, a szelekció semmit nem tesz a variabilitás nélkül, és ez részben az élőlényt érő hatásoktól függ. Sokszor a természetet megszemélyesítettem, mivel nehéz volt ezt a kettős értelmet elkerülni; de én természet alatt csak a természeti törvények összesített hatásait és termékeit, ill. törvények alatt csakis bizonyos események sorozatát értem.1
Az élőlényeknek azonban szaporodniuk is kell, nemcsak túlélni; máskülönben a tulajdonságaik nem adódnak át az utódoknak. Bármi, ami egy élőlény sikeres szaporodásához hozzájárul (vagyis képes olyan utódokat létrehozni, amelyek maguk is továbbélnek, majd szaporodnak) az állat „fitnesz”-ét (rátermettség) erősíti, vagyis a faj képes lesz egy bizonyos környezetben tartósan megmaradni. Az, hogy a környezet milyen mértékben határozza meg a fitnesz értékét, arról sokat vitatkoznak, de ez volt Darwin alapvető ötlete.

Amint olvastuk, Darwin szintén egyetértett Spencernek a „legrátermettebbek túléléséről” alkotott fogalmával, de sok mai evolucionista nem kedveli ezt a fogalmat, mivel az embereket arra befolyásolja, hogy a „legnagyobb”, „leggyorsabb”, vagy „legerősebb” fogalmakban gondolkozzanak, noha ezek a tulajdonságok nem mindig teszik az embert képessé, hogy életképes utódokat hozzanak létre. A „legfittebb” az, aki definíció szerint a legtöbb életképes utódot hozza létre. Akinek legtöbb a gyereke, az nyer! Sok zavar van ekörül a téma körül, egészen Darwinig visszamenően. A fenti idézet előtt épp azt írja „Bizonyára mondták, hogy az egész természet hadban áll; a legerősebbek végül kitartanak, míg a gyengék elesnek.” A zűrzavar ellenére, a biológusok a „természetes szelekció” fogalmát a differenciált reprodukcióra használják. Ez egy nagyon fontos megkülönböztetés.
Darwin a Fajok eredetében a természetes szelekciót hozta fel egyetlen mechanizmusként, hogy a földön meglévő sokféle életforma létrejöttét megmagyarázza; egyetlen eredeti létformából (avagy néhány létformából, amit Darwin megengedett). Nem ismerte sem a genetikát, sem a mutációkat, sem a DNS-beli alapjukat (lásd 2. fejezetet). Azt vetette fel, hogy kis változások mindig is vannak, és azok, amelyek biztosították a túlélést, megmaradtak, ezzel a fajt egy egészen másik fajjá alakítva (ha elég idő van rá).
A természetes szelekció bizonyítékát egyben az evolúció bizonyítékaként kezelik. Mivel az élőlények természetes szelekció útján sokszor képesek a környezetükhöz alkalmazkodni, bőven akad a természetes szelekcióra példa, így egyfolytában azzal bombáznak minket, hogy az evolúció „egyfolytában zajlik”. De valóban ez lenne az evolúció?
Az evolúció „speciális”, illetve „általános” elmélete
Mi az evolúció? „Időbeli változás”, vagy „minden faj közös eredete”? Miközben Darwin azzal próbálkozott, hogy ezt a két fogalmat összekösse, Darwin elmélete új fajok kialakulását feltételezte (speciáció), noha nemigen magyarázta meg azt, hogy új fajok hogyan is alakultak ki (és hogy hogyan történik, az ma is vita tárgya).
Később írok arról, hogy a „faj” fogalmát hogyan határozták meg. Most legyen elég annyi, hogy a speciáció egyszerűen azzal foglakozik, hogy egy nyúlfaj hogyan ered olyan ősi nyúlfajokból, amelyekkel együtt már nem alkot szaporodási közösséget. Ez teljesen eltér attól, mikor egy új fajt úgy közelítünk meg, mint a mikrobák és az ember közti egyik láncszemet.
 Darwin azt feltételezte, hogy a fajok közti sokféleség végtelen, így tehát a természetes szelekció megfelelően hosszú idő alatt egy mikrobát mongúzzá tudott alakítani. Feltételezte, hogy a kutya- és házigalamb-fajták, vagy a különböző vadon élő pintyek2 csőrméretének sokfélesége azt a változási sort mutatta be, amelyet a végtelenségig vissza lehetett vezetni, így az nemcsak a különböző pintyfajokat, hanem még a pintyek, galambok, kutyák és minden más faj eredetét is megmagyarázza. Nem foglalkozott az ember eredetével, csak 12 évvel később, „Az ember származása” című művében, valamint a „Szexuális szelekció” című könyvében, feltehetően azért, mert ha az ember eredetéről írt volna „A fajok eredeté”-ben, sokkal kevesebb esélye lett volna arra, hogy elfogadják.
Darwin azt feltételezte, hogy a fajok közti sokféleség végtelen, így tehát a természetes szelekció megfelelően hosszú idő alatt egy mikrobát mongúzzá tudott alakítani. Feltételezte, hogy a kutya- és házigalamb-fajták, vagy a különböző vadon élő pintyek2 csőrméretének sokfélesége azt a változási sort mutatta be, amelyet a végtelenségig vissza lehetett vezetni, így az nemcsak a különböző pintyfajokat, hanem még a pintyek, galambok, kutyák és minden más faj eredetét is megmagyarázza. Nem foglalkozott az ember eredetével, csak 12 évvel később, „Az ember származása” című művében, valamint a „Szexuális szelekció” című könyvében, feltehetően azért, mert ha az ember eredetéről írt volna „A fajok eredeté”-ben, sokkal kevesebb esélye lett volna arra, hogy elfogadják.
Meglehetősen nagy ugrás a meglévő jellemzőn belüli sokféleségről (mint a rövidebb, vékonyabb, hosszabb, illetve vastagabb csőr) a csőrök eredetére, vagy rögtön a pintyek, madarak, hüllők, emlősök, illetve minden más faj eredetére következtetni. Hogyan magyarázza a kutyafajták közötti változatosság a kutyák (farkasok) eredetét? Itt alapvető logikai hiba van, ami nagyon jól megvilágítja az evolúció Achilles-inát.
Valóban, a híres brit professzor G. A. Kerkut, a gerinctelen állatok szakértője, megkülönbözteti az „evolúció speciális elméletét” (speciáció) az „evolúció általános elméletétől” (ami minden élőlény közös eredetét jelenti). Arról érvel, hogy az utóbbi, az általános elmélet csupán spekuláció:
Az „Evolúció Általános Elméletét” a bizonyítékok nem támasztják alá kellőképpen ahhoz, hogy az elméletet munkahipotézisnél többnek tartsuk. Nem világos, hogy azok a változások, amelyek új fajokat hoznak létre, ugyanolyan természetűek, mint azok, amelyek új rendszertani törzseket hoznak létre [ezek az élőlények nagyobb rendszertani egységei, amelyből 80 létezik, és amibe a mikrobák is beletartoznak]. A választ majd jövőbeni kutatómunka fogja feltárni és nem azon dogmatikai kijelentések, hogy az Evolúció Általános Elmélete csak azért is igaz, mert nincs más, ami kielégítően megmagyarázza a jelenségeket.3
Mellesleg, Kerkut az élet eredetét is belefoglalta az Evolúció Általános Elméletébe. Miért van az, hogy manapság annyian nem akarják belevenni az élet eredetének kérdését az evolúció definíciójába? Lásd a 3. fejezetet.
Ma tudjuk, hogy miért nem lehet a fajok közti kis változtatásokat (Speciális Elmélet) minden élőlény változatosságának eredetére is kiterjeszteni (Általános Elmélet). Az a fajta megfigyelhető sokféleség, amit az evolucionisták „evolúció”-nak neveznek, attól van, hogy meglévő genetikai információt (alléleket) összekevernek, vagy egy olyan véletlen folyamat, ami majdnem mindig degeneratív (elfajulást okozó) és információvesztéssel jár. Ennek ellenére, a mikroorganizmusoktól az emberig tartó evolúciónak egyfolytában új, komplex információ-gazdag génekre van szüksége, hogy izomsejteket, csontokat, idegeket, tollakat hozzon létre, ahol korábban azok nem léteztek. Darwin nem is sejtette, hogy mit igényelnek ekkora nagymérvű változások, és hogy a modern biológia elképesztő komplexitást tárt fel, ami szemben áll azzal a gondolattal, hogy ez az idő során felhalmozódó kis változásoktól van.
1980 novemberében a világ egyes vezető evolucionistái „történelmi” konferenciát tartottak Chicago természettudományi múzeumában. Roger Lewin ezt írta a konferenciáról a Science folyóirat lapjaiban:
A chicagói konferencia központi kérdése az volt, hogy a mikroevolúció mechanizmusai kiterjeszthetők-e a makroevolúcióra. Megkockáztatva, hogy esetleg egyes konferencia résztvevők álláspontját megsértjük, a válasz egy egyértelmű nem.4
Francisco Ayala, aki akkoriban a California Egyetemen genetika professzora volt, kijelentette, hogy most arról van meggyőződve, hogy „a kis változások nem halmozódnak fel”.5
Ennek ellenére sok evolucionista kitartóan semmibe veszi ezt az állítást. Azaz, még mindig azt az elképzelést terjesztik, hogy a
„Nagy Változás = Kis Változás x Több Millió Év”. Ez egy olyan logikai átverés, hogy mást adnak, mint amit kínálnak. Olyan ez, mintha azt mondanám, hogy „mivel egy tehén át tud ugrani egy kerítést, csak idő és gyakorlás kérdése, hogy majd a holdat is átugorja”.
Egyes tanárok ehhez hasonló trükköt használnak, hogy a diákjaikkal, akik ellenállnak az evolúció általános elméletének, elfogadtassák, hogy az evolúció egyenlő a változással. „Tessék, itt a példa a változásra, tehát az evolúció tény!”
Egy másik hamis meghatározása az evolúciónak az, amit „allél gyakoriság változásnak” hívnak. Az allélgyakoriságok tényleg megváltoznak, de ez nem magyarázza meg a gének eredetét (a génvariánsok pedig az allélok), amit a makroevolúció taglal, hanem csak a meglévő allélok gyakorisága változik.
John Endler, elsőrangú evolucionista, valamint az USA Nemzeti Tudományos Akadémia választott tagja, így beszél erről:
Az evolúció nem más, mint bármilyen irányú változás vagy felhalmozódó változás az élőlények vagy populációk tulajdonságaiban sok generáción keresztül … Ebbe beletartozik az allélok eredete, valamint az allélok elterjedése, vagy azok variánsai, tulajdonságbeli értékei, vagy karakterállapotai.6
Sok evolucionista úgy beszél, mintha az evolúció csakis az utóbbiról szólna, és maguknak a tulajdonságoknak az eredetét kihagyja a tárgyalásból. Darwin írásaiból ez a tény nyilvánvalóan kimaradt, így a mai evolucionista gondolkodásban is elterjedt, de ettől függetlenül hamis.
Az evolúció nem pusztán „változás”. Nem azonos a tulajdonságok gyakoriságainak megváltozásával (allél frekvencia) egy populációban (speciális elmélet). Teljesen új bélyegek eredetét is magába kell, hogy foglalja, amelyek nemcsak meglévő allélok megváltozásai (általános elmélet), és ez az, ahol Darwin, majd sokan utána kudarcot vallottak.
A természetes szelekció nem evolúció
 Sok magas rangú evolucionista úgy beszél a darwinizmusról/evolúcióról és a természetes szelekcióról mintha azonosak lennének. Például Dr. Richard Dawkins olyan kísérletekről számol be, amelyekben a természetes szelekciót a guppik színalakulásával (egyik szerepe az álcázás a ragadozókkal szemben, másrészt a hímnemű egyedek gazdag színvilágukkal vonzzák a nőstényeket) jellemzik, és úgy állítják be, mint „az evolúció csodálatos példája, ami a szemünk előtt zajlik”.7
Sok magas rangú evolucionista úgy beszél a darwinizmusról/evolúcióról és a természetes szelekcióról mintha azonosak lennének. Például Dr. Richard Dawkins olyan kísérletekről számol be, amelyekben a természetes szelekciót a guppik színalakulásával (egyik szerepe az álcázás a ragadozókkal szemben, másrészt a hímnemű egyedek gazdag színvilágukkal vonzzák a nőstényeket) jellemzik, és úgy állítják be, mint „az evolúció csodálatos példája, ami a szemünk előtt zajlik”.7
A fent idézett Dr. John Endler az, aki ezt a kutatást végezte a guppikról. Egyetértene Dawkinssal, hogy „ez az evolúció csodálatos példája”?8 A vadonban történő természetes szelekció című könyvében, amit 1986-ban írt, Endler azt állította, hogy a kettő nem ugyanaz:
A természetes szelekció nem ugyanaz, mint az evolúció, noha a kettő közeli kapcsolatban áll egymással.9 A természetes szelekció elég gyakori a vadon élő populációkban ahhoz, hogy sok fajban megfigyeljék, és az erős szelekció nem is olyan ritka, mint ahogyan korábban feltételezték; így a természetes szelekció valószínűleg fontos szerepet játszik az evolúcióban. Ennek ellenére a természetes szelekció nem magyarázza az új variánsok eredetét, csakis a gyakoriságukban végbemenő változásokat.10
Így tehát a természetes szelekció befolyásolhatja a tulajdonságok kombinációinak mintázatát, de nem magyarázza a tulajdonságok eredetének mechanizmusát. Erről érintőlegesen írtak a következő szerzők: Fisher (1930), Simpson (1944) és Rensch (1959), de azóta szinte semmilyen figyelmet nem kapott. A téma mindenképp további kutatást érdemelne.11
Jegyezzük meg, hogy Fisher, Simpson és Rensch igen magas rangú evolucionisták voltak. Fisher például a modern evolúciós szintézis egyik megalapozója.
A guppi kutatást illetően, noha a természetes szelekció a színes és színtelen guppik relatív gyakoriságát meg tudja magyarázni, a szexuális szelekció12 (ahol a nőstények az élénkszínű hímeket kedvelik) és a predáció között egyensúlyozva (ahol a halványszínű halak vannak előnyben), de nem magyarázza a színek eredetét. Noha egyes mutációk egy meglévő színre ható génben befolyásolni tudják a guppi kinézetét, ez még nem magyarázza magának a génnek az eredetét. Ez egy fontos különbség, amivel maga Darwin és az utána következő evolucionisták következetesen keveset foglalkoztak.
Dawkins és társai tényleg nem tudják, hogy a természetes szelekció nem egyezik az evolúcióval, noha magas rangú evolucionisták (mint akire maga Dawkins is hivatkozik, hogy állítását ezzel alátámasztja) világosan beszéltek erről?
Darwin példáját követve az evolucionisták mégiscsak arról szeretnek beszélni, hogy a természetes szelekció az egy kreatív erő, noha nem hoz létre semmit. Pusztán arra képes, hogy a kevésbé rátermett egyedeket eltüntesse, semhogy a rátermettséget létrehozza. A természetes szelekció nem ugyanaz, mint az evolúció. „Az erősek túlélése” (a gyengék eltűnése) nem tudja megmagyarázni azt, hogy az erősek hogyan jönnek létre.
A természetes szelekció nem Darwin ötlete volt
Ha figyelembe vesszük, hogy a természetes szelekció mennyire nyilvánvaló tény, nem meglepő, hogy ez az elképzelés nem Darwinnal kezdődött.

Maga Carl von Linné (1707–1778), a híres svéd taxonómus, aki a fajok elnevezésének és osztályozásának ma is használt rendszerét megalapozta, és akit a „rendszertan atyjának” hívnak, láthatóan megértette a természetes szelekció fogalmát, ill. az életben maradásért folytatott küzdelemben elfoglalt szerepét. A kaliforniai egyetem őslénytani múzeumának honlapján Linnéről ezt írják:
Linné megfigyelte az életben maradásért folytatott küzdelmet – amit a Természet „vágódeszkájának” nevezett, valamint a „mindenki ellen vívott harc”-nak hívott. Ennek ellenére a küzdelmet és a versengést szükségesnek tartotta a természeti rend fenntartásában, ami az isteni rend része volt.13
James Hutton, skót deista 1794-ben (egy 2000 oldalas, 3 kötetes publikálatlan műnek az egyik fejezetében14) azt írta a kutyákról, hogy azoknál, melyeknél a túléléshez csak a gyorsaság számít, a lassabb kutyák elpusztulnak, és a gyorsabbak maradnak fenn. Ám hogyha a jó szaglásra „nagyobb szüksége volt az állatnak az életben maradásért”, akkor a „fajtán belül, ugyanazon az elven olyan kutyák jönnek létre, amelyek a prédájukat a kifinomult szaglásukkal találják meg, és nem a gyorsaságukkal kapják el áldozatukat. Azt írta, hogy ugyanez a „variálódás elve” hat „minden növényfajra, függetlenül attól, hogy az az erdőben vagy a réten nő”.15
Hutton világosan a természetes szelekcióról írt, ha nem is ezt a kifejezést használta (azt a fogalmat csak 1859-ban találták ki, amikor megjelent a Fajok Eredete).
Egy skót-amerikai orvos, William Wells (1757–1817), 1813-ban a természetes szelekció fogalmának egy fajtáját írta le. Afrika egyes lakóiról azt mondta, hogy:
…jobban alkalmazkodnának a betegségekkel szemben, mint más csoportok. Ez a rassz pedig ennek következtében elszaporodna, míg a többiek elfogynának.16
Azt feltételezte, hogy ez az életerős rassz sötét bőrű lenne, és hogy:
…mivel a legsötétebb bőr alkalmazkodott legjobban az adott éghajlathoz, idővel a legelterjedtebb, ha nem az egyetlen rasszá válna abban az országban ahonnan származott.17
Patrick Matthew (1790-1874) skót gyümölcstermelő 1831-ben adta ki „A hajófa és a fakitermelés” című könyvét, amelynek függelékében megemlítette a természetes kiválasztódást. Matthew később nyilvánosan kijelentette, hogy megelőzte Charles Darwin-t, sőt könyvének lapjain „a természetes kiválasztódás eredete felfedezőjének”18 nevezi önmagát. Darwin tagadta, hogy tudott volna Matthew munkásságáról és rámutatott, hogy ez egy jelentéktelen könyv függelékében jelent meg. Mindennek ellenére, ha maga Darwin nem is tudott erről a konkrét példáról, ez azt bizonyítja, hogy hasonló gondolatok már ismertek voltak a viktoriánus társadalomban 1859 előtt.
Pearson rámutatott, hogy Wells, Matthew és Darwin egyaránt Edinburgh-ban, ebben az egyetemi városban éltek, „amely tudományos klubjairól és társaságairól volt híres”, emellett Hutton szülővárosa is volt.19 Valóban nagyon különös lenne, ha Darwin nem tudott volna ezekről az előzetes ötletekről, és így akár tudatosan, akár akaratlanul, de az elmélete az említett társasági körökben forgó filozófiai elmélkedések olvasztótégelyében alakult ki. A lényeg az, hogy a természetes kiválasztódás ötlete egyszerűen nem Darwintól származik, hanem ez abban az időben egy általánosan elterjedt gondolat volt, legalábbis bizonyos körökben.
Valószínűleg Edward Blyth (1810-1873) gyakorolta a legnagyobb hatást Darwinra. Blyth, az angol vegyész és zoológus három fontos cikket írt a természetes kiválasztódásról, ezek a cikkek 1835 és 1837 között jelentek meg a Természettudományi Szemlében, tehát abban az időszakban, amikor Darwin még Föld körüli úton volt a Beagle fedélzetén.
A Természettudományi Szemle abban az időben az egyik vezető zoológiai folyóirat volt, amelyben Darwin barátai, tehát Henslow, Jenyns és Lyell szintén publikáltak cikkeket.
Halála előtt Loren Eiseley (1907-1977) az antropológia és a tudománytörténet Benjamin Franklin-díjas professzora volt a Pennsylvaniai Egyetemen. Eiseley évtizedeken keresztül foglalkozott a fajok eredete Darwinnak tulajdonított elméletével. 1979-ben megjelent utolsó könyvében ezt írta: „Darwin művének egyik vezető tételét – a létért folytatott küzdelem, a változások, a természetes kiválasztódás és a nemi kiválasztódás – teljes mértékben kifejtette Blyth az 1835. évi tanulmányában.”20 Miért nem ismerte el Darwin Blyth munkásságát a természetes kiválasztódás tekintetében? Russell Grigg véleménye szerint21:
1. Blyth keresztyén volt, és manapság a „speciális teremtés” hívének neveznék [legalábbis korai munkássága alapján]. Az állatok színének évszakonkénti változásairól (például, amikor a havasi nyúl színe télen fehérré változik) Blyth azt állította, hogy ezek „a tervezés feltűnő esetei, amelyek egyértelműen és kényszerítő erővel tanúsítják egy mindentudó és hatalmas eredeti forrás létezését”. Azt is mondta, hogy az állatok „a természetfeletti bölcsesség bizonyítékai, mert ösztöneiket csak egy mindentudó Teremtőtől kaphatták”.22
2. Blyth a természetes kiválasztódás elvét helyesen olyan mechanizmusnak tekintette, amely kiszűri a populációból a beteg, öreg és életképtelen egyedeket, vagyis fenntartó erőnek tekintette, amely rögzíti a teremtett faj eredeti állapotát. Az olyan [speciális] teremtés hívei, mint például Edward Blyth [akivel kapcsolatban léteznek arra utaló bizonyítékok, hogy A fajok eredete című könyv megjelenése után ő is magáévá tette Darwinnak a korlátlan változásokkal kapcsolatos elméletét, az ember és a majom közös származását is ide számítva], és William Paley angol teológus a természetes kiválasztódást olyan selejtezési folyamatnak tekintették, ahol a tulajdonságok már mind a kezdetkor megvannak, mielőtt a kiválasztás elkezdődne.
Csak néhány olyan személyt említettünk meg, akik Darwin előtt már írtak a természetes kiválasztódásról. Nem feledkezhetünk meg Alfred Russel Wallace (1823-1913) nevéről sem. Mialatt az Indonéz szigetvilágban élt, önállóan kidolgozott egy evolúciós elméletet, amely csaknem teljesen megegyezett Darwin elméletével, ide számítva a természetes kiválasztódás teljesen kialakult elméletét is. 1858-ban elküldte Darwinnak „A változatok azon hajlamáról, hogy tetszőleges mértékben eltérjenek az eredeti típustól” című kéziratának egy példányát, amelyben leírja mindazt, amit az evolúció darwini elméleteként ismerünk. Sokan úgy vélik, hogy Darwin és barátai, Charles Lyell és Joseph Hooker méltánytalanul bántak vele, és mindent megtettek azért, hogy megszerezzék Darwinnak az elsőbbséget a londoni Linné Társaságnál.

Ami az evolúciót illeti (ami tehát nem azonos a természetes kiválasztódással) már a kereszténységet megelőző korszakokban is sokan felvetették, hogy minden élőlény valamiféle ősi előd(ök)ből fejlődött ki.23
Darwin keresztényellenes nagyapja, Erasmus Darwin is kiadott 1794-ben egy kétkötetes értekezést az evolúcióról „Zoonómia” címmel. Családi címerükön ez a latin jelmondat szerepel: E conchis omnia, vagyis „minden a kagylóból ered”. Tehát Darwin nagyapja nem egyszerűen hitt az evolúcióban, hanem ezt bele is illesztette a család nyilvánosság felé mutatott arcába, vagyis a család címerébe!
Már Darwin előtt is sok írás megjelent Európában különféle evolúciós ötletekről, melyek megkísérelték megmagyarázni mindennek az eredetét isteni teremtés nélkül.24 A Teremtő Isten lerázásának vágya mélyen beágyazódott az emberi jellembe, és ez nem Darwintól származik.
Darwin előfutárai, például Hutton és Matthew, a természetes kiválasztódást az életképességet konzerváló és fenntartó tényezőnek tekintették, amely változásokat csak bizonyos korlátok között képes előidézni. Darwin azonban teremtő erőként hívta segítségül a természetes kiválasztódást, és ezzel próbálta megmagyarázni minden élőlény eredetével kapcsolatos evolúciós nézeteit. Ebben különbözött előfutáraitól, és pontosan ebben tévedett, mert a természetes kiválasztódás nem rendelkezik teremtő erővel.
A természetes kiválasztódás nem a teremtés cáfolata
A Kaliforniai Egyetem Paleontológiai Múzeumának honlapján olvasható cikk ezt mondja Linnéről:
Korai éveiben Linné úgy vélte, hogy a fajok nem egyszerűen valóságosak, hanem változtathatatlanok. Saját szavaival élve: Unitas in omni specie ordinem ducit (A fajok változtathatatlansága a rend feltétele [a természetben]). Később Linné megfigyelte, hogy a különféle növényfajok keresztezhetők egymással, és ezáltal új fajnak tűnő létformákat lehet létrehozni. Ezért elhagyta azt az elgondolást, hogy a fajok állandóak és változtathatatlanok, majd felvetette, hogy néhány, vagy talán a legtöbb létező faj a világ teremtése után állt elő, hibridizáció útján. Mivel azzal kísérletezett, hogyan lehet idegen növényeket meghonosítani Svédországban, Linné azt is feltételezte, hogy a növényfajok megváltoztathatók úgy, ha betelepíti azokat. Életének vége felé Linné azt vizsgálta, hogy génuszok között is létezik-e a keresztezés lehetősége, és azt feltételezte, hogy hibridizáció útján esetleg új génuszok is létrejöhetnek.
Vajon Linné evolucionista volt? Az igaz, hogy eltávolodott korábbi véleményétől, amely szerint a fajok változtathatatlanok, és az is igaz, hogy keresztezéssel létrehoztak már új növényfajokat, egyes esetekben állatfajokat is. Linné azonban az új fajok létrehozását nem tekintette végtelen és korlátlan lehetőségnek. Bármilyen új fajok jöttek is létre az eredeti fajokból, az Édenkert eredeti fajaiból, ezek továbbra is Isten teremtő tervének részét alkották, mert mindig jelen volt a lehetőség a létezésükre. A végtelen evolúció gondolata, amely nem áll szükségszerűen az isteni terv irányítása alatt, és amelynek nincs előre meghatározott célja, soha meg sem fordult Linné fejében, és ez a gondolat nagyon megdöbbentő lett volna a számára.25
Más szavakkal: úgy tűnik, Linné eredetileg úgy vélte, hogy a fajok voltak a teremtett típusok, de később felismerte, hogy a teremtett típusok hasonló fajok, sőt akár teljes nemzetségek is lehettek. A „fajok állandóságának” elképzelése (vagyis hogy új fajok nem lehetségesek) olyan ősi tekintélyektől származik, mint például Arisztotelész; és a Linnét megelőző korokban az ősi tekintélyekre való hivatkozás nagyon fontos volt. A Biblia latin fordítása (Vulgata), amelyben a „faj” (min) héber szót fajnak fordították, szintén ezt az elgondolást támogatta, jóllehet ezt a szót nemzetségnek vagy más ma ismert egyéb rendszertani kategóriának is lehetne fordítani. Tehát Linné 1700-as években tett megállapításai ellenére az 1800-as évek közepén az volt az általánosan elterjedt nézet, hogy a fajok változtathatatlanok, és az egyes fajok csak ugyanolyan fajokat képesek létrehozni, és soha nem alakulhatnak ki belőlük más fajok. A deizmus volt az uralkodó nézet az értelmiségi körökben (pl. Lyell), ami együtt járt egyfajta személytelen természetfeletti „eredeti okba” vetett hittel, de ez nem volt azonos azzal a Teremtővel és Megváltóval, aki megmutatta magát a történelemben és a Bibliában. Ezen vélemény szerint „a világegyetem nagy mérnöke” teremtette a különféle fajokat, többé-kevésbé a mai állapotukban. Ezen kívül a Darwin napjaiban regnáló egyház már kapitulált a hosszú korszakok és azok következményei előtt és tagadta a Noé napjaiban bekövetkezett özönvizet.26 Ezért nem lehet meglepő az élőlények természetéről kialakított nem biblikusnak tekinthető elképzelések elfogadása.
 A „fajok állandósága” kellemes szalmabáb-célt biztosított Darwin számára. Meglehetősen nyilvánvaló volt, hogy a Galápagos-szigeteki pintyek a kontinens pintyeitől származtak, és ezt követően alakultak ki belőlük a nyilvánvaló módon eltérő „fajok” a szigeteken (jóllehet Darwin akkor nem ismerte fel, hogy ezek pintyek voltak, és csak később eszmélt rá ennek a jelentőségére, amikor John Gould pintyként azonosította ezeket a madarakat). Abrakadabra! Megdőlt a fajok állandósága. Ám soha senki nem végzett kísérleteket annak bizonyítására, hogy ezek a pintyfajok keresztezhetők-e egymással. A közelmúltban elvégzett egyes tanulmányok azt mutatják, hogy a kereszteződés természetes módon megtörténik27, tehát Darwin pintyei még biológiai fajnak sem tekinthetők! És Linné felismerte volna, hogy ugyanahhoz a teremtett fajhoz tartoznak.
A „fajok állandósága” kellemes szalmabáb-célt biztosított Darwin számára. Meglehetősen nyilvánvaló volt, hogy a Galápagos-szigeteki pintyek a kontinens pintyeitől származtak, és ezt követően alakultak ki belőlük a nyilvánvaló módon eltérő „fajok” a szigeteken (jóllehet Darwin akkor nem ismerte fel, hogy ezek pintyek voltak, és csak később eszmélt rá ennek a jelentőségére, amikor John Gould pintyként azonosította ezeket a madarakat). Abrakadabra! Megdőlt a fajok állandósága. Ám soha senki nem végzett kísérleteket annak bizonyítására, hogy ezek a pintyfajok keresztezhetők-e egymással. A közelmúltban elvégzett egyes tanulmányok azt mutatják, hogy a kereszteződés természetes módon megtörténik27, tehát Darwin pintyei még biológiai fajnak sem tekinthetők! És Linné felismerte volna, hogy ugyanahhoz a teremtett fajhoz tartoznak.
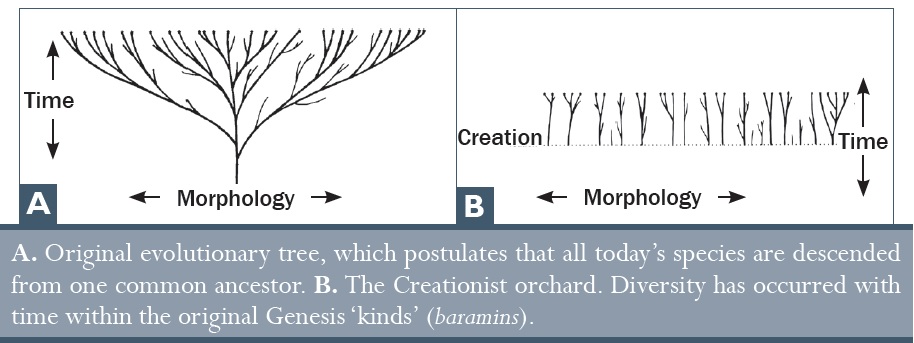
Ezzel a nézettel Linné előre jelezte a mai kreacionista biológusok véleményét: az élő szervezetek képesek alkalmazkodni és új „fajokat” létrehozni, de ennek a folyamatnak a hatóköre korlátozott és a tapasztalható változások jellege nem ad magyarázatot az élő szervezetek alapvetően eltérő típusainak eredetére. Amit látunk, az csak az élőlényekbe teremtett alkalmazkodási- és diverzifikációs képesség kibontakozása a teremtett típusok korlátain belül, hogy a Föld különféle ökológiai térségeit kitölthesse az élet. A természetes kiválasztódás ennek a folyamatnak a részét alkotja, amint ezt a kreacionista Linné és Blyth már régen felismerte. Ha tehát a természetes kiválasztódás a kreacionista modell része, hogyan lehet a teremtés gondolatának cáfolata, vagy pedig az evolúció bizonyítéka a teremtéssel szemben? Ha az evolúció és a teremtés is igényt tart ugyanarra a területre, akkor valami mással kell érvelni. Így tehát az érvelés eltolódik az evolúció speciális elveitől (a fajokon belüli viszonylag jelentéktelen változásoktól), amelyek nyilvánvalóak, egyértelműek és megfigyelhetőek, az evolúció általános elvei felé (minden élőlény sokféleségének eredete), amely az élet biokémiai bonyolultságával kapcsolatos mai felfogásunk szerint napjainkban sokkal, de sokkal nehezebben hihető, mint Darwin korában volt.
Mi a „faj”?
 Az oxfordi szótár szerint a biológusok a fajt úgy határozzák meg, mint „egymáshoz hasonló élőlények közössége, amelyek képesek géneket kicserélni vagy egymással párosodni”. Ezt sok biológus elfogadja, beleértve a kreacionistákat is. Más szavakkal, ha két élőlény egymással tud párosodni, akkor szigorúan ugyanabba a (biológiai) fajba tartoznak. Ennek ellenére, mivel ilyen hibridizációs kísérleteket nehéz elvégezni, a fajneveket gyakran a kinézetük (morfológiájuk) alapján adják, így nem konzisztensek a párzást megakadályozó határokkal (sokkal több fajnév létezik, mint biológiai faj).
Az oxfordi szótár szerint a biológusok a fajt úgy határozzák meg, mint „egymáshoz hasonló élőlények közössége, amelyek képesek géneket kicserélni vagy egymással párosodni”. Ezt sok biológus elfogadja, beleértve a kreacionistákat is. Más szavakkal, ha két élőlény egymással tud párosodni, akkor szigorúan ugyanabba a (biológiai) fajba tartoznak. Ennek ellenére, mivel ilyen hibridizációs kísérleteket nehéz elvégezni, a fajneveket gyakran a kinézetük (morfológiájuk) alapján adják, így nem konzisztensek a párzást megakadályozó határokkal (sokkal több fajnév létezik, mint biológiai faj).
Amikor azonban fosszilis maradványokról van szó, a párosodást nem lehet vizsgálni. Az évmilliókban, hosszú idősíkokban való gondolkodás azt is befolyásolja, hogy a geológusok, illetve az őslénytanosok milyen neveket adnak a fosszilis fajoknak. Amikor olyan kövületet találnak, amelyet több millió évesnek datálnak, de ugyanakkor ma is élő fajra hasonlít, az átívelő evolúcióba vetett hitük arra vezeti őket, hogy kizárják annak a lehetőségét, hogy talán ugyanarról a fajról van szó. Több ezernyi „élő kövület” van, amely ugyanúgy néz ki, mint a mai fajok, de szinte mindig más nemzetségbe sorolják, nemhogy más fajba.28 Ez a félrenevezés azt a hamis benyomást kelti az élőlényekről, mintha sokat változnának, holott csak keveset, vagy egyáltalán nem is változtak semmit. Ez kiválóan mutatja , hogy az ember alapfeltevése meghatározza azt, hogy hogyan értelmezi a „tényeket”.

Olyan kövületek, amelyeket más kontinensen is megtalálnak, szintén más nevet kapnak, így növelve a zűrzavart. Képzeld el, hogy a különböző kutyafajták ma nem élnének. Ha a paleontológusok a nagy dán dogtól illetve a csivavától találnának fosszíliákat, érdekes lenne, hogy milyen neveket adnának nekik. Majdnem biztosan más fajnevet adnának nekik, sőt lehet, hogy más nemzetségbe is sorolnák őket. Ugyanabba a családba tennék őket? Ez a gondolat azt húzza alá, hogy nem az a fontos az evolúcióban, hogy mennyi a változás, hanem hogy milyen típusú ez a változás. Miközben a dán dog és a csivava nagyon eltérően néznek ki, tudhatjuk, hogy ezek a különbségek csak fokozatbeli különbségek: ezek meglévő tulajdonságok módosulatai, és nem teljesen új bélyegek. Nem jelentek meg tollak vagy csőrök, vagy más „nem farkas” tulajdonságok. Ilyen tulajdonságok létrejönnek, ha elég idő van rájuk? Minél többet tudunk meg ezen tulajdonságok bonyolult genetikai, illetve biokémiai hátteréről, annál nehezebb hinni az evolúcióban.
Az 1. Mózes 1:11-ben a Biblia arról beszél, hogy Isten a növényeket teremti meg: „Azután monda Isten: Hajtson a föld gyenge füvet, maghozó füvet, gyümölcsfát, amely gyümölcsöt hozzon az ő neme szerint, amelyben legyen neki magva e földön. És úgy lőn.” (Károli)
Ezt a formulát, „az ő neme szerint”, egymás után használják (tízszer) a különböző teremtett típusokra Mózes első könyvének első fejezetében. Ez egy alapvető biológiai elvet húz alá, amit mindenki egyfolytában megfigyelhet, és amit gyerekkorunk óta értünk: az élőlények a saját nemüket hozzák létre: kutyák kutyákat hoznak létre, macskák macskákat, gyümölcsfák pedig gyümölcsfákat, stb. Senki más nem figyelte meg ennek az ellenkezőjét (a 4. fejezetben lesz majd szó olyan bizonyítékokról, amelyek az időbeli változás mellett, illetve ellen szólnak), de az evolúció azon a tévhiten alapul, hogy egy alapvető élőlénytípus valami nagyon mássá alakult át (például az, hogy a giliszták emberekké alakulnak, ahogyan egy kiemelkedő evolucionista beszélt róla a New Scientist (Új Tudós) című lapban29). A speciációt (a teremtett típusokon belüli változásokat) az általános evolúció elméletének bizonyítékaként értelmezik, de amint ezt korábban bemutattuk, ez elfogadott része a teremtés-modellnek is.
A hibridizációs események utalnak arra, hogy a mai élőlények hogyan alakulhattak ki az eredeti teremtett típusokból. A teremtett típusok bibliai meghatározása alapján a kreacionisták azt a következtetést vonják le, hogy ha két faj tud hibridizálódni valódi megtermékenyítéssel, akkor ez a két egyed ugyanabból a típusból származik.30 Továbbá, ha bármely egyed a kettő közül egy harmadikkal is tud hibridizálódni, akkor mindhárman ugyanabba a típusba tartoznak.31 Fordítva, ha két egyed nem tud közös hibridet létrehozni, akkor nem feltétlenül igaz, hogy nem ugyanabba a típusba tartoznak, mivel a mutációk degenerációhoz vezethetnek, mint pl. a kromoszomális átrendeződések, amelyek párzási akadályokat háríthatnak olyan egyedek közé, amelyek máskülönben azonosak lennének.
 1985-ben a hawaii Sea Life Park-ban bálnaborjú született egy hím kis kardszárnyúdelfintől (Pseudorca crassidens) és egy nőstény palackorrú delfintől (Tursiops truncatus).32 Az utód nagyon meglepte a park dolgozóit, mivel a két szülőfaj nagyon másképpen nézett ki. Itt olyan hibrid keletkezett, amely a Delphinidae családon belül (delfinek, kis kardszárnyúdelfinek és kardszárnyúdelfinek), de eltérő nemzetséghez tartozó szülőktől eredt. Mivel az ivadék ebben az esetben termékeny (a hibrid nőstény azóta borjút is szült), ez a két nemzetség valóban definíció szerint „politípusos” biológiai faj. Más nemzetségek, amelyek ugyanebbe a csoportba tartoznak, jobban hasonlítanak egymásra, mint az a két egyed, amely ezt az utódot létrehozta Hawaii-on, amely erősen arra utal, hogy a tizenkét élő nemzetség ugyanabból a teremtett típusból származhatott. Ilyen események arra emlékeztetnek, hogy a modern rendszertani kategóriák mesterségesek, amelyeket eredetileg Linné hozott létre, de amit ma arra használnak, hogy az általános múltbeli evolúciós eseményeket meghatározzák. Nem éppen az a fajta egzakt tudomány, mint aminek láttatják.
1985-ben a hawaii Sea Life Park-ban bálnaborjú született egy hím kis kardszárnyúdelfintől (Pseudorca crassidens) és egy nőstény palackorrú delfintől (Tursiops truncatus).32 Az utód nagyon meglepte a park dolgozóit, mivel a két szülőfaj nagyon másképpen nézett ki. Itt olyan hibrid keletkezett, amely a Delphinidae családon belül (delfinek, kis kardszárnyúdelfinek és kardszárnyúdelfinek), de eltérő nemzetséghez tartozó szülőktől eredt. Mivel az ivadék ebben az esetben termékeny (a hibrid nőstény azóta borjút is szült), ez a két nemzetség valóban definíció szerint „politípusos” biológiai faj. Más nemzetségek, amelyek ugyanebbe a csoportba tartoznak, jobban hasonlítanak egymásra, mint az a két egyed, amely ezt az utódot létrehozta Hawaii-on, amely erősen arra utal, hogy a tizenkét élő nemzetség ugyanabból a teremtett típusból származhatott. Ilyen események arra emlékeztetnek, hogy a modern rendszertani kategóriák mesterségesek, amelyeket eredetileg Linné hozott létre, de amit ma arra használnak, hogy az általános múltbeli evolúciós eseményeket meghatározzák. Nem éppen az a fajta egzakt tudomány, mint aminek láttatják.
A kreacionista biológus Dr. Frank Marsh ezekről a fogalmakról az 1940-es években sokat írt.
A zoológus Wayne Frair ezt mondta ezzel kapcsolatban33:
Marsh meggyőzően érvelt amellett, hogy a tudományos bizonyítékok az egymástól elkülönült típusok elképzelését támasztják alá, amelyek sokféle változatot hoztak létre, fosszilis, illetve ma is élő formákban. Az ebben a könyvben lévő anyag nagyon fontos volt számomra, hogy a „teremtett típusok” fogalmát elfogadjam.
Marsh a „baramin” fogalmat alkalmazta a teremtett típusra (a héber bara szóból, ami azt jelenti, hogy „teremteni”, és a min szóból, ami azt jelenti, hogy „fajta” vagy „típus”), továbbá különböző kritériumokat hozott létre, hogy meghatározza ezen típusok utódait. A „baraminológia” így a teremtett típusok rendszertani elemzése. Például Dr. Jean Lightner állatorvos azt vizsgálta, hogy a juhok illetve a kecskék ugyanabba a baraminba tartoznak-e. Lightner így határozta meg az eredményeit:
… a juhok (Ovis aries) és a kecskék (Capra hircus) egyetlen monobaraminba tartoznak (egy olyan csoport, amely ugyanabba a típusba tartozik). További hibridizációs adatok azt mutatják, hogy más fajok, amelyek az Ovis, Capra, Ammotragus, Hemitragus, és valószínűleg a Rupicapra nemzetségekbe tartoznak, ugyanebbe a monobaraminba esnek. Egy állítólagos hibrid, amit egy juh és egy európai őz között hoztak létre, azt mutatja, hogy ez a monobaramin akár több kérődző családot is magába foglalhat; egy jobban dokumentált esetre lenne szükség, mielőtt ilyen következtetéseket vonhatnánk le. Az a fajta variáció, amely ebben a monobaraminban létezik, és amely adaptív változásokat is magában foglal, azt mutatja, hogy a mutációk és a kromoszomális átrendeződések sokat alakítottak a ma élő fajokon.34
Ezen új „fajok” kialakulása nem okoz problémát a teremtés számára
Világos, hogy az új fajok kialakulása része a teremtés modellnek. Új biológiai fajok létrejöttét megfigyelték már. Így például „új fajok” létrejöttek a Drosophila nemzetségben, amelyet olyan gyakran vizsgálnak az egyetemi genetikai laboratóriumokban. Egy gyakori mechanizmus az az inverzió, amikor találomra egy kromoszóma részlet átrendeződik (ez egy fajta mutáció), ahol a kromoszómának egy része irányt vált. Ez a „génáramlást” meggátolja azon muslicák között, amelyek rendelkeznek az inverzióval, illetve azok között, ahol nincsen meg ez az inverzió, így a két csoport genetikailag eltérő csoportba rendeződik. Ez azt eredményezi, hogy egymástól egyre jobban elhatárolódnak, noha még képesek hibrideket létrehozni.35 Az ilyen eseményeket „evolúció”-nak nevezik, azonban genetikai értelemben új fajok nem jönnek létre, ami ahhoz kellene, hogy egy légyből valami mást hozzunk létre – itt pusztán a kromoszómákon lévő gének fizikai átrendeződéséről van szó. Ennek tehát semmi köze az egysejtűektől az emberig tartó metafizikai folyamat hiedelméhez, vagy az evolúció általános elméletéhez, ahol pusztán természetbeni folyamatok hozták volna létre a ma látható sokféle életformát egyetlen közös ősből.
A bibliai teremtésmodell azt is magába foglalja, hogy a tüdővel lélegző, földön élő életformák Noé idejében az özönvíztől elpusztultak, és minden állattípusból egy-egy pár, helyenként hét-hét pár élte túl a bárkán. Azok az állatok, amelyek a mai ökológiai niche-eket betöltik, azokból az állatokból származnak, amelyek a bárkán fennmaradtak. Azt a következtetést vonjuk le, hogy rendkívül sok variáció történt abban a 4500 évben, ami az özönvíz óta eltelt, illetve hogy az adaptáció és a speciáció időnként rendkívül gyorsan mentek végbe. Valóban, elég bizonyíték áll rendelkezésre arról, hogy gyorsan is végbemehet a speciáció, ami az evolucionistákat mindig meglepi, akik eleve „hosszú korszakok”-ból indulnak ki.36

Például Trinidadban dolgozó kutatók guppikat (Poecilia reticulata) helyeztek át egy vízeséses területről, ami tele volt ragadozóhalakkal, a vízesések fölé, ahol addig nem voltak guppik, és csak egy predátorfaj fordult elő, amely viszont csak a kisebb méretű guppikra vadászott, így a nagyobb méretű guppik fennmaradtak. Az áthelyezett guppik ivadékai úgy alkalmazkodtak az új körülményekhez, hogy nagyobbra nőttek, később lettek ivarérettek, illetve kevesebb, de nagyobb ivadékaik voltak.37
Ezen változások sebessége meglepte az evolucionistákat, mivel az évmillió évek alatt zajló változásokba vetett hitük jóval hosszabb időszakot követelt az adaptációhoz. Egy evolucionista azt mondta:
A guppik mindössze négy év alatt alkalmazkodtak a környezetükhöz – ez a változás pedig 10.000-10.000.000-szoros sebességű, mint a kövületekből megállapítható átlagos változás mértéke.38
És sok más ehhez hasonló példa is van.
A fajkeletkezés folyamata nem támogatja az evolúciót
Semmiféle fajkeletkezési folyamat nem létezik, ami azt bizonyítaná, hogy a fajkeletkezés újabb fajokhoz vezetne. Például, ha a fenti guppi megfigyelésekre hivatkozunk, egy evolucionista genetikus azt mondta, hogy „amennyire én tudom, ezek még mindig guppik”.39
A biológusok a bölcsőszájúhal-félék családjába tartozó halakat sokat kutatták, amelyek három kontinensen megtalálhatók, illetve sok közülük akváriumi díszhal. Nagyon változatos a kinézetük és sokszor úgy mutatják be őket, mint az evolúció folyamatának élő példáját. Azonban a teremtéshívő zoológus Dr. Arthur Jones a doktori disszertációját e halak változatosságáról írta, de ezzel épp a teremtésbe vetett hitét igazolta. Dr. Jones a következőket írja:
A kutatás évei csodálatosak voltak. A fajok változatosságát vizsgálva azt találtam, hogy a bölcsőszájúhal-félék tévedhetetlenül természetes csoport, egy teremtett típus. Minél többet foglalkoztam ezekkel a halakkal, annál jobban megértettem, miből áll a bölcsőszájúhal-félék egyedi volta abban, hogy egyre jobban elkülönültek azoktól a hozzájuk „hasonló” halaktól, amelyeket tanulmányoztam. A konferenciák alatt történt beszélgetések és az irodalmi kutatásaim azt igazolták, hogy ez az állatrendszertan területén dolgozó összes szakember véleménye. A különálló típusok tényleg léteznek, és a szakemberek ezt tudják is. A fejlődésbiológiai tanulmányok azt mutatják, hogy a bölcsőszájúhal-félék óriási változatosságát (több mint ezer faj) valóban a viszonylag kevés karakterállapot végtelen permutációja okozta: négy szín, körülbelül tíz alapvető színmintázat, stb. Ugyanazok a karakterek (vagy karakter mintázatok) „véletlenszerűen” megjelentek a különböző fajokban. A variáció mintázatai „modulárisak” avagy „mozaikosak” voltak; evolúciós leszármazási vonalak sehol nem voltak megtalálhatók. Ez a fajta adaptív variáció elég gyorsan meg tud történni (mivel olyan genetikai anyaggal dolgozik, ami már eleve ott van), és néhány esetben a bölcsőszájúhal-félék szétterjedése (a geológiailag „fiatal” tavakban) valóban mérhető volt (az evolucionisták által), pusztán néhány ezer évre datálva. Tágabb értelemben a fosszíliák nem adtak bizonyítékot az evolucionisták kezébe. Minden hal, élő vagy fosszília formájában, alapvetően teremtett típusokba tartozik; az „összekötő láncszemek” nyilvánvalóan hiányoznak.40

A fajkeletkezés mögött húzódó genetikai folyamatok közé tartozik a meglévő genetikai információk rendszerezése és újrakombinálódása, ahogyan ezt Arthur Jones az előbb megmutatta. A genetikai aberrációk (mint a kromoszómák inverziója) szaporodási határokat hozhatnak létre és ezzel segíthetnek a fajkeletkezésben.
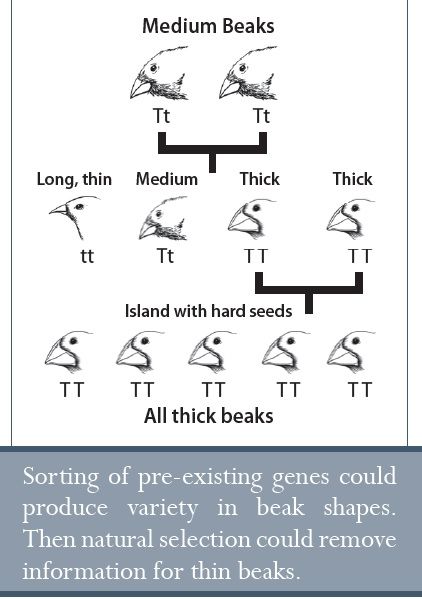
Továbbá az adott környezeti niche-hez történő adaptációval járó fajkeletkezésben az alkalmazkodni képtelen egyedek kiszelektálódnak. A korábbi példánkban, a rövid szőrű farkasok a sarkkörhöz közel kiválasztódnak, azt eredményezve, hogy a rövid szőrzetért felelős gének kiesnek a populációból. Ezért a fajkeletkezés által történő adaptáció valóban „kiszór” valamennyi genetikai anyagot a populációból. Amint a rövid szőrzetért felelős gének kiesnek a populációból, a farkasok nem tudnak a meleg éghajlathoz alkalmazkodni, így ők a hideg éghajlathoz vannak már „berendezkedve”.
Ez az evolúciós fajkeletkezés problémáira vet fényt. Ha a természetes szelekción keresztül történő adaptáció egyfolytában kigyomlálja a genetikai információt (variáció), a fajok egyre jobban idomulnak a saját környezetükhöz. Ez a specializálódás pedig annak vet gátat, hogy visszaalakuljanak, ha a környezet megváltozik. Bizonyos értelemben javára válna egy fajnak, ha általános módon tudna válaszolni a környezeti változásokra, ám a szelekció a fajkeletkezés irányába hajtja őket. Ezért a fajok olyan niche-ekben határolódnak el az idők során, ahonnan nem könnyű elmenekülni. Ezt a jegesmedve esetében láthatjuk, amely a „normális” medve ősöktől származott41, de ami nem nagyon tudna boldogulni, ha a Sarkvidék felmelegedne, és a jég elolvadna. A fajok adaptáción alapuló gyors megváltozása, amit részben a természetes szelekció vezérel, szépen beleilleszkedik a teremtés-modellbe.
Senki nem figyelt még meg olyan fajkeletkezést, ami egy újabb biokémiai folyamatot igényelne. Továbbá, senki sem hozott létre egy működőképes modellt ilyen változásokra. A fajkeletkezés a populációkban meglévő genetikai információ átrendeződésével vagy elvesztésével történik. A fajkeletkezés nem támasztja alá az evolúció általános elméletét.
A természetes szelekció nem hoz létre újabb genetikai információt
A természetes szelekció csakis a meglévő genetikai információra tud hatni, ami meghatározza a populációban megtalálható tulajdonságokat. Nem tud információt létrehozni. A legtöbb evolucionista ezt érti is, de nem tudja jól kommunikálni a nyilvánosság felé.
Az evolúció modern felfogása szerint (neo-darwini szintézis) a mutációk hoznak létre újabb információt (pl. a fehérjékért felelős specifikációk). Sokféle mutáció létezik, de mindegyikük a DNS betűsorrendjében történő véletlen megváltozással jár együtt. Röviden, a mutáció az egyetlen létező folyamat arra, hogy az élőlényekben lévő hatalmas mennyiségű információt létrehozza; vagyis a tollak, a csontok, az izmok, az idegek, a haj, vagy a hemoglobin specifikációját, vagy az emberi magzat alig ismert fejlődéséhez szükséges fejlődési állapotok összehangolását. Ez a fajta specifikáció mind hiányzott az első feltételezett mikróbából, az egyetemes közös ősből (universal common ancestor – UCA), de egy sor mutáció elvileg mindegyiket hozzáadta, apró lépésenként, minden egyes változással annyi előnyt adva, hogy a természetes szelekció „lássa” és rögzítse (szakkifejezéssel „fixálja”) a populációban.
Ezek a szekvenciális mutációk továbbá új biokémiai folyamatokat is létre kellett hozzanak, beleértve számtalan enzimet (vagyis fehérjéket), vagy olyan nanogépeket, amelyek több fehérjekomponensből állnak (mint az ATP-szintáz42, az RNS polimeráz, a helikáz, illetve a giráz).
A probléma abban áll, hogy ezeknek a biokémiai utaknak vagy nanogépeknek egyik komponense sem tudna a véletlen folyamán (mutációk révén) előállni, nem is beszélve az összes komponensről; mindegyikre egyszerre van szükség, hogy ezek a gépek működhessenek. Valóban, sok biokémiai folyamat toxint termel, ha még nem teljes (ez húzódik meg a patogén baktériumok toxicitása mögött, ahol egy mutált gén egy enzimet kiiktat, ami annak az anyagnak a felhalmozódásához vezet, ami a hiányzó enzim szubsztrátja lenne).
Ahhoz, hogy egy prokarióta szervezetet (mint pl. egy baktériumot) eukariótává alakítsuk (mint pl. az élesztő, a növények vagy az állatok), a mutációknak ki kell találniuk a kinezin transzport rendszert, amellyel az eukarióták rendelkeznek, ami több integrált komponensből áll, ami mind szükséges a rendszer működéséhez.43
Ezek kikerülhetetlen akadályok a véletlen változások (mutációk) számára, hogy létrehozzanak, ill. módosítsanak már meglévő genetikai rendszereket.
A mutációk szükségesek az evolúcióhoz
Mennyi információt kell a mutációknak a genomhoz adni, hogy egy kezdetleges mikróbát emberré alakítsanak? A legegyszerűbb mikróba genomja körülbelül 500,000 nukleotidból („betűből”) áll.44 Egy embernek három milliárd betűje van. Tehát ahhoz, hogy az előző élőlényt az utóbbivá alakítsuk, ahhoz a mutációknak létre kell hozniuk majdnem három milliárd nukleotidnyi információt/specifikációt. Egyedül a fehérjék szintjén egy egyszerű baktériumnak néhány száz fehérjéje van, az embernek azonban több mint 100,000. Továbbá az embereknek az összes olyan instrukcióval is rendelkezniük kell, amelyek az embrionális fejlődést vezérlik (amik az evolúciós megközelítés miatt, számtalan biológus által „hulladék”-nek gondolt DNS 98%-ában találhatók, ld. 2. fejezet).
Amikor a mutációkat legelőször felfedezték, az evolucionisták azt az új információt létrehozó mechanizmusnak vélték, ami az egyszerű életformáktól az emberig tartó fejlődésért felelős. Ez az elgondolás lett az „új” darwinizmus, vagyis a „modern szintézis” egyik fontos eleme, ami a harmincas-negyvenes években kezdett formálódni.
Az ionsugárzás illetve bizonyos vegyszerek mutációkat indukáltak, így a növénytermesztők a „mutációs nemesítés” pályájára léptek, abban a reményben, hogy mindenféle csodálatos új növényfajtát illetve fajt tudnak majd létrehozni, ami segít majd az emberiségnek. Ebben az evolúció gyorsított változatát látták. Nagy pénzösszegeket költöttek ezekre a programokra, de az eredmények annyira kiábrándítóak voltak, hogy ma kevés ilyen programot tartanak fent a fejlett országokban.
Néhány hasznos mutációt azonban találtak, mint ami a törpe növekedést okozza, (ami abban segíti a növényt, hogy ne dőljön össze a sok mag súlya alatt), törhetetlen maghüvely (ami abban segít, hogy a magok betakarításkor ne a földre essenek), alacsony fitát tartalmú kukorica (amit a tehenek is meg tudnak enni), vagy a változatos színek a dísznövényekben, mint például a krizantém esetében.45 Ha azonban ezeket molekula-szinten vizsgáljuk, azt látjuk, hogy valami mindig összetört ahhoz, hogy ezek az új tulajdonságok kialakuljanak, semmint, hogy keletkezett volna valami. Ez világos a törpenövekedés, illetve a törhetetlen maghüvely esetében. Az alacsony fitát tartalmú kukorica esetében a mutációk megsértették azt a metabolikus utat, ami a fitát létrehozza. A virág színvariációk esete annyira nem világos, de ez a többféle pigment kombinálódásából ered. Egy olyan mutáció, ami az egyik pigmentet kiüti és más színt eredményez, gazdaságilag is értékes lehet.46
A muslicák (Drosophila spp.) a genetikai tanulmányok kedvenc célfajai, különösen a hallgatói projektek számára. Számtalan Drosophila mutációs kísérletet végeztek el vegyszeres illetve sugárzásos módszerekkel, ami mindenféle defektes legyet hozott létre, de semmi nem vezetett ahhoz, hogy bebizonyítsák, hogy ezek a mutációk újabb genetikai specifikációt tudnak létrehozni, ami a legyeket bármi mássá átalakítaná.

Az evolúciót legfeljebb a mikróbákban tudjuk megfigyelni
Ha az evolúció bárhol tetten érhető, az csakis a mikróbák köre. Sok baktériumfajt először az 1800-as évek végén fedeztek fel és neveztek el, de még ma is ugyanazokkal a jellegzetességekkel rendelkeznek.
A baktériumoknál fennáll egy olyan tendencia, ami a felfelé tartó evolúcióval szembemegy – sok új gén létrehozása helyett a baktériumok törlik a felesleget.47 Ez különösen az obligát paraziták tenyészeteiben látható, ahol sok gén eredeti formájában nem szükséges a túléléshez, így ezek törlődnek és kiesnek a genomból.
1988-ban Dr. Richard Lenski a Michigan állami egyetemen 12 E. coli kultúrát hozott létre, majd ezeket a laborjában húsz éven át fenntartotta, generációról generációra (kitartásáért dicséretet érdemel). A táptalaj valamennyi glükózt tartalmazott, de még több citrátot, így mire a mikróbák elfogyasztották a glükózt, csak akkor tudtak tovább növekedni, ha a citrát felhasználását valahogyan kifejlesztik.
Lenski azt várta, hogy majd megfigyeli a felgyorsított evolúciót. Ez egy megfelelő elvárás volt olyasvalakitől, aki hisz az evolúcióban, mivel a baktériumok gyorsan tudnak szaporodni, és így nagy populációik lehetnek. Emellett magasabb mutációs rátájuk is lehet, olyan élőlényeknél, amelyek nagyobb genommal is rendelkeznek, mint a gerincesek, mint amilyenek mi is vagyunk. A neo-darwinizmus szerint ez mind ahhoz vezet, hogy bizonyosan nagyléptékű evolúciót valós időben megfigyeljenek (ahelyett hogy a távoli múltban elképzeljék). A rövid generációs időt figyelembe véve, 20 év mintegy 44.000 generációt jelent, ami néhány millió évnek felel meg emberi léptékkel. Ugyanakkor, az embernél sokkal kevesebb az evolúció lehetősége a kisebb populációból adódó kevesebb mutációs lehetőség miatt; ill. a sokkal nagyobb és komplexebb genom nem tud ugyanakkora mutációs rátát fenntartani hibázási katasztrófa nélkül (pl. kihalás); mindamellett a szexuális szaporodás révén a jótékony mutáció jó eséllyel nem is adódik tovább.
Sok eredménytelen év után Lenski felhagyott a „laboratóriumi evolúcióval”, és inkább az evolúciót egy Avida nevű program segítségével akarta modellezni. Lenskinek valóban oka volt arra, hogy elveszítse a reményt. Kiszámolta, hogy minden lehetséges egyszerű mutációnak már többször végbe kellett volna mennie, de egyetlen új tulajdonság sem jelent meg. Egyszer csak mégis nagy felhajtással bejelentették, hogy a 12 baktériumtörzs közül egy megszerezte azon képességet, hogy a citrátot mint alapanyagot használja a 31.500. generáció után.
Csakhogy az E. coli tud citrátot bontani anaerób körülmények között. Mindössze annyi kell hozzá, hogy aerób körülmények között is felhasználja, hogy megszakítsuk azt a mechanizmust, ami oxigén jelenlétében a citrát felvételt gátolja; ezt én 2008-ban már felvetettem.48 Sokkal könnyebb összetörni, semmint létrehozzni valamit. És ahogy azt már láttuk, az evolúciónak nagyon nehezére is esik valamit létrehozni. Lenski laborja ezek után megtalálta azt a mutációt, ami felelős ezért az új tulajdonságért, amit 2012-ben közöltek.49 A citrát egy transzporter fehérje. Ezt a fehérjét a citrát transzporter gén, a citT kódolja, ami normális körülmények között, oxigén jelenlétében ki van kapcsolva. Nagyon közel a citT génhez vannak más olyan gének, amelyeknek a promótere oxigén jelenlétében ezeket a géneket bekapcsolja. Egyetlen mutáció azt okozta, hogy a promótert egy olyan helyre másolta, ahol a citT gént bekapcsolta, úgy, hogy a citrát transzporter fehérje oxigén jelenlétében is termelődött. További mutációk megduplázták a citT gént, így a baktériumok a citrát transzporterből többet termeltek, és még több citrátot vettek fel.
Mint teremtéskutató biológus, arra gyanakodtam, hogy valami itt eltört; az pedig a citT felvételének a mechanizmusa oxigén jelenlétében. A mutációk nem hoztak létre újabb gént vagy promótert; egyszerűen azt másolták, ami már jelen volt a baktérium genomjában. Ebben a folyamatban egy olyan baktérium jött létre, ami már többé nem tudja a citT gént kikapcsolni oxigén jelenlétében (megj.: ha ezek a baktériumok kikerülnének a laboratóriumi körülmények közül, a természetben nem tudnának túlélni, mivel a transzporter fehérje termelése nagy pazarlás, amikor nincs rá szükség).
Ez megegyezik Dr. Michael Behe analízisével, aki egy másik mikróbát vett szemügyre, aminek nagyméretű populációi vannak: a malária parazitáját, a Plasmodiumot.50 Többféle maláriaellenes gyógyszer ellen már rezisztens, de az emberek maguk is kifejlesztettek ellenállást (pl. a sarlósejtes anémia, illetve a talasszémia). Behe azt mutatta ki, hogy az adaptáció minden esetében, mind a Plasmodiumban, mind az emberben, a génállományban valami összetört, nem pedig egy újabb, komplexebb tulajdonság jött létre. Például a Plasmodiumban a klorokin rezisztenciájáért egy transzport fehérjében történő hiba a felelős, ami a mérget az élőlény vakuólumába (sejtnedvüreg) szállítja be.
Behe ezt a jelenséget az első világháborús árkokban történő háborúhoz hasonlítja, ahol a védők a saját hídjukat vagy útjait azért robbantják fel, hogy a támadókat ezzel lelassítsák. Ez valójában nem fegyverkezési verseny, mivel ott az egymással szembenálló erők mindig újabb fegyvereket találnak ki; azok a természeti folyamatok („evolúció”), amelyek a Plasmodiumban illetve az emberekben végbemennek, nem járnak újabb „fegyver” létrehozásával.
Behe a Plasmodiumban lévő pirimetamin rezisztenciát is vizsgálta, a DDT rezisztenciát a szúnyogokban, illetve a warfarin rezisztenciát a patkányokban. Mindhárom esetben a mutációk mindig valamit összetörtek, hogy a rezisztenciát létrehozzák.
Az emberi betegségeket okozó mutációkat tartalmazó adatbázisa a mutációk okozta károkat világítja meg. 2011 októberében az Online Mendelian Inheritance in Man (az Emberi Mendeli Öröklődés) nevű adatbázis51 2.665 olyan gént sorolt fel, amelyekben egy vagy több mutáció ismert betegséghez vezet. A közegészségügyi szakértők attól óvnak, hogy az emberek kitegyék magukat a mutagének illetve az ionsugárzás okozta hatásoknak, mivel azok betegségeket okoznak és nem szuperhősöket hoznak létre (akiknek a csodaképességei elvileg a mutációktól keletkeznek).
A mutációs tenyésztés illetve a mikróbák mutációi ismét azt hangsúlyozzák, hogy a random változások (mutációk) nem hoznak létre újabb genetikai anyagot, hogy egyszerűbb élőlényeket komplexebbekké alakítsanak, amelyek újabb tulajdonságokkal rendelkeznek, amelyekhez újabb DNS specifikáció szükséges (vadonatúj gének és ellenőrzőrendszerük).
Azt látjuk, hogy mindezek ellenére a mutáció az egyetlen mechanizmus az evolucionisták számára, hogy megmagyarázhassák a bioszférában megtalálható hatalmas mennyiségű genetikai információt. Időbeli változás? Igen. Olyan változás, ami miatt a mikróbák mikrobiológusokká alakultak? Dehogy! Egyszerűen nem jön ki a számítás.
A természetes szelekció a megőrzésre törekszik
Darwin azt gondolta, hogy a természetes szelekció a nagyon kis variációt is érzékeli. Ez azonban naiv elgondolás volt. A valóságban a természetes szelekció csak az erős hatásokat tudja detektálni, amelyek például halált okoznak, vagy amelyek a halált megelőzik. Kimura azt fedezte fel, hogy a mutációk túlnyomó többsége túl kicsi hatást fejtenek ki ahhoz, hogy a természetes szelekció érzékelni tudná; a mutációknak létezett egy olyan tartománya, melyek láthatatlanok voltak a természetes szelekció számára. Ezek a mutációk benne voltak „Kimura dobozában”.
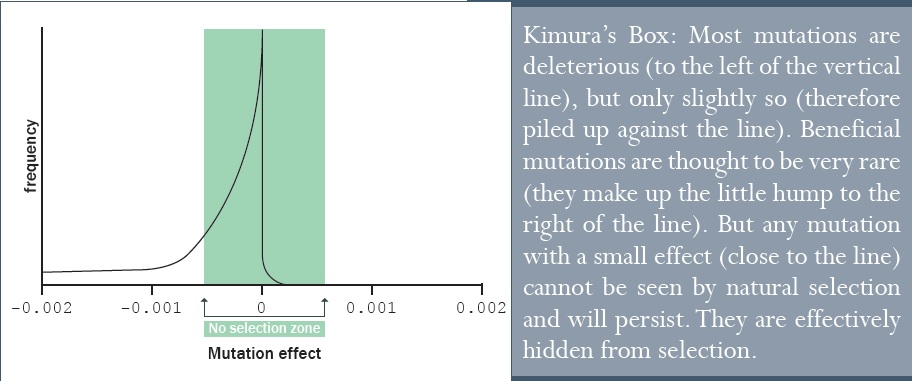
Egy másik evolucionista, J.B.S. Haldane, az 1950-es években azt számolta ki, hogy az új tulajdonságoknak legalább 10%-os szelekciós előnyt kell hozniuk ahhoz, hogy a populációban rögzülhessenek (a 10%-os előny azt jelenti, hogy az adott tulajdonsággal rendelkező egyedek 10%-kal több túlélő utódot hoznak létre azokkal az egyedekkel szemben, amelyek nem rendelkeznek azzal az előnyös tulajdonsággal).52
Az evolucionisták a populációgenetika illetve az evolúciós folyamat és annak időtartama alapján azt feltételezték, hogy az élőlényekben lévő mutációs ráta meglehetősen alacsony, körülbelül egy mutáció egyedenként egy generáción belül, vagy akár ennél is kevesebb. Amint felismerték, hogy a legtöbb mutáció káros, azt is gyanították, hogy nem lehet sok belőlük, különben a természetes szelekció nem tudná a káros mutációk túlsúlyát megszüntetni; egyfajta mutációs leállás történne, különösen a több millió éves idősíkban.
A közelmúltban a mutációs ráta mérésekor azt találták, hogy az 50-szer magasabb, mint amit addig az evolucionista ideológia szerint vártak. Ez hatalmas problémát okoz az evolúcióelmélet számára. Dr. John Sanford, a Cornell egyetem nyugdíjazott genetikusa (jelenleg tiszteletbeli professzor) és a génpuska felfedezője53, kimutatta, hogy a mutáció gyors üteme valamint a legtöbb mutáció enyhén káros volta (ami által Kimura dobozába esnek) azt jelenti, hogy ezek az enyhén káros mutációk láthatatlanok a természetes szelekció számára, és így az emberekben illetve más élőlényekben felgyülemlenek. Ez a folyamat kérlelhetetlenül elpusztít, és nem épít minket. Mi a kihalás felé haladunk, minden más komplex élőlénnyel együtt.54
Sanford azt a kérdést teszi fel, hogy ha a legtöbb, információval ellátott nukleotid (a DNS betűi) önmagában keveset ad hozzá a genomhoz, akkor hogyan került oda, és hogy maradt ott olyan sokáig? A természetes szelekció nem látja őket egyenként; csak a teljes fitnesz értéket látja (ami a túlélő- és szaporodási képességet jelenti), és az egyetlen nukleotid hozzáadott értéke annyira kevés, hogy láthatatlan; az összes többi nukleotid „zajában” elvész. Dr. Sanford az evolúció problémáit így összegezte53:
1. a mutációk sokkal gyorsabban keletkeznek annál, semhogy a szelekció hathatna rájuk;
2. a mutációk túlnyomó többsége olyan kis hatással rendelkezik, hogy a természetes szelekció számára nem láthatók;
3. a biológiai zaj és a „szerencsések túlélése” a szelekciónál erősebb hatások;
4. a káros mutációk sokszor jó hatású mutációkkal vannak összekapcsolva, így azokat nem tudják szétválasztani az öröklődés során (hogy a rosszakat eltüntessék, illetve a jókat megtartsák). Ennek az az eredménye, hogy az összes komplex genom romlik.
A tájékozott evolucionisták tudnak ezekről a problémákról, és ezért a „szinergisztikus episztázis” elméletét hozták létre (ahol több mutáció együttes hatása több mint az egyedi mutációk összege).55 Sanford azonban kimutatta, hogy ettől a probléma csak nagyobb lenne. Dr. Tomoko Ohta, evolucionista és Kimura egyik legjobb tanítványa, akivel sok közös cikket is írt, a „populációgenetika királynője” lett. Az amerikai tudományos akadémia tiszteletbeli tagja. Sanford egyik munkatársa a szinergisztikus episztázisról kérdezte, és egyetértettek abban, hogy az elmélet csak tovább bonyolítja a kérdést55. Sanford és munkatársai számítógépes szimulációkat is folytattak egy kifinomult populációgenetikai modellel („Mendel Könyvelője”)56, hogy ezt bizonyítsák.
Egy olyan folyamat, ami egy genomot lassanként degradál (ezzel egyidejűleg a „genetikai entrópiát” növeli), nem tud jobb élőlényt létrehozni hosszú távon. Sanford analízise lerombolja az evolúciós paradigmát. Nehéz elképzelni, hogyan lehet megvédeni ezzel a kritikai elemzéssel szemben. Emellett egy nagytekintélyű genetikustól egy ilyen állítást nem lehet könnyen lesöpörni.
Ennek ellenére az evolucionisták egyfolytában az adaptív mutációk által történő „fejlődés” példáit próbálják felmutatni nekünk, abban a reményben, hogy elhisszük. Ám az általuk felhozott példák mind egy szerv vagy tulajdonság elvesztéséről szólnak: látás-vesztés a barlangi halaknál57 és szalamandráknál58, a szárny elvesztése szeles szigeten élő bogaraknál59, az enzimtermelés kontrolljának megszűnése vagy a fehérjecsatorna károsodása antibiotikum-rezisztenciát előidézve60, vagy bizonyos halak defektív génjei, amelyek révén meg tudnak élni PCB-vel szennyezett vizekben.61 Más szavakkal, „összetört” élőlényeket mutatnak nekünk, mint az adaptív mutáció és a természetes szelekció példáit.
Tüskés pikók
A tüskés pikókat (Gasterosteus spp.) is az evolúciót bizonyító csoportnak tartják, mert a természetes szelekcióra, sőt, egy adaptív (előnyös) mutációra is példát mutatnak, de ismét csak azt bizonyítják, hogy nem létezik olyan folyamat, melyben a haltól a halászig jutnánk (általános evolúcióelmélet).
A tüskés pikók két alakban vannak jelen. Van egy tengeri és egy édesvízi alak. A sósvízi alaknak a testén tüskék vannak és számos csontlapocskával, ún. páncéllal rendelkezik. Ezek megvédik a ragadozóktól. Az édesvízi alak nagy morfológiai változatosságot mutat, de általában rövidek a háti és hasi tüskék és lényegesen kevesebb a csontlapocskák száma; vagy nincs is páncéljuk.62
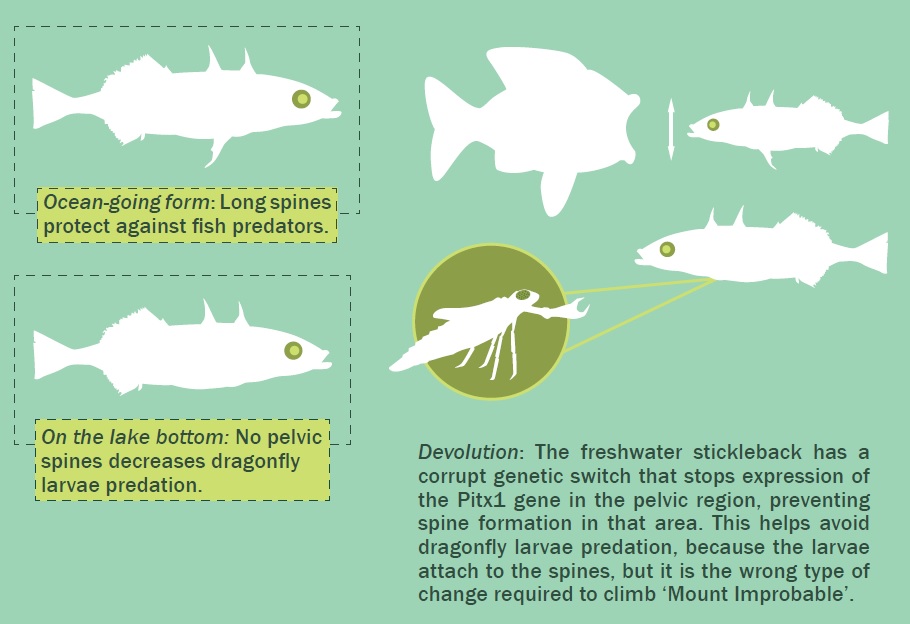
Úgy tartják, hogy az édesvízi alak a sósvízi alakból fejlődött ki, és ez lehetséges is. Sőt, belefér a teremtésmodellbe, az özönvíz utáni benépesítés és diverzifikáció kapcsán.
A tengeri alak a folyókon keresztül az édesvízi tavakig vándorol, ahol a két alak szaporodhat is egymással, ami arra utal, hogy ugyanabba a teremtett típusba (baraminba) tartoznak.
A biológusok megfigyelték, hogy amikor sósvízi tüskés pikókat édesvízi tavakba telepítenek, több év után a csontlapocskák mennyisége, illetve a tüskék mérete lecsökken. A páncél és a tüske több tényező miatt is előnytelen a tavakban: kevesebb a ragadozó, a páncél növesztése a kalciumhiányos vizekben sok erőforrást igényel, illetve a tó fenekén leselkedő ragadozó szitakötő lárvák a tüskénél ragadják meg a felettük átúszó pikókat.
Az édesvízi környezetben azok a pikók, amelyeknek kevesebb csontlapocskájuk van és nincs hasi tüskéjük nagyobb túlélési előnnyel rendelkeznek, vagyis a természetes szelekció abba az irányba hat, hogy a halaknak kevesebb páncélja és tüskéje legyen.
Mi áll a változások mögött? Az evolúció valamilyen újabb tulajdonságot fejlesztett volna ki? A genetikusok olyan mutáció kiváltotta „genetikai kapcsolót” fedeztek fel, amely befolyásolja a Pitx gén kifejeződését. A hasi oldalon ez a mutáció akadályozza meg, hogy kialakuljon a tüske. Másutt egy sor csontképlet kialakulását szabályozza63, így nemcsak a páncélra, hanem az állkapocs alakjára és még a kopoltyút védő csontokra is hat. Más szavakkal, a pikók édesvízi alakjai egy genetikai kapcsoló miatt jöttek létre, ami több jellemzőt is ki tud iktatni.
Dr. Jerry Coyne (University of Chicago), aki az evolúcióelmélet nagy propagálója, a pikók genetikai kapcsolóját elemezve őszintén bevallotta, hogy „ez a genetikai tulajdonságok elvesztésével jár együtt, és nem jár evolúciós újdonságok létrejöttével.”64 Ezeket a különbségeket mégis változatlanul az evolúció bizonyítékaként kezelik. A 2009-es Darwin Év kapcsán a Nature szakfolyóirat a pikókat a „15 evolúciós ékszer” egyikeként mutatta be, Sean Carroll, egy magas tekintélyű evolucionista pedig „az evolúció egyik legmeggyőzőbb bizonyítékaként” tartja számon.65
Itt a természetes szelekció, az adaptív mutáció, sőt a fajkeletkezés egyik példájával állunk szemben, de ebben az esetben sincs bizonyíték arra, hogy a tüskék, illetve a páncél újonnan keletkeztek volna mutációk, illetve természetes szelekció révén.
Még a mutált genetikai kapcsoló is megjelent a sósvízi pikókban, ám kisebb számban.
A fenti folyamat fordítottját is megfigyelték: amikor a Washington tavat (Seattle közelében) megtisztították a szennyeződéstől, és így a tóban a látási viszonyok rohamosan javultak, a kevesebb páncéllal rendelkező pikók az ott honos pisztrángok (Oncorhynchus clarkii) számára könnyebb prédát jelentettek, így a teljes páncélzattal rendelkező pikók aránya 6%-ról 49%-ra emelkedett öt évtized alatt. Ezt felgyorsított, fordított irányú evolúciónak mondták. Ennek ellenére, semmi újat nem hozott létre ez az evolúció. A teljes páncélzatért felelős gének már jelen voltak a populációban (eredetileg 6%-ban), és a természetes szelekció egyszerűen ezt a gyakoriságot megnövelte.66 Ez nem más, mint tipikus félremagyarázás; „az evolúció csupán a gén-gyakoriság megváltozása”.
Itt semmi nem támasztja alá az evolúció általános elméletét, mely szerint újabb gének alakulnak ki, és nem egy meglévő genetikai kapcsoló széttörését (ami már létezett is), vagy a meglévő gének gyakoriságának megváltozását.
A genetikai kapcsolókat az adaptáció vezérléséhez teremtették?
A kreacionista kutató állatorvos Dr. Jean Lightner azt feltételezte, hogy a genetikai kapcsolók, mint amik a pikók esetében is lehetővé teszik az adaptációt, olyan módon jöttek létre, hogy mutálódhatnak anélkül, hogy más tulajdonságokat károsan érintenének. Talált egy fehérjét, ami többfunkciós kapcsolóként működik, és az állatokban a sokféle szőrszínt hozza létre.
Mivel ez a fehérje csak a szőr színét befolyásolja, jól tolerálja a mutációt. Amikor normálisan működik, a fehérje az eumelaninnak nevezett sötét színű pigment termelését kapcsolja be. Továbbá, egy halvány sárgás-pirosas pigmentet is létrehoz.
A mutációk a kapcsolót úgy tudják kiiktatni, hogy a sötét pigment termelését nem indítja el. Ez a golden retriever sárga színezetét hozza létre, például. Más esetben a kapcsoló „bekapcsolva marad”, így az eumelanin túltermelése révén sötét színt eredményez.
Dr. Lightner hozzáteszi:
A Teremtő ezt a különleges kapcsológént azzal a képességgel hozhatta létre, hogy oly módon változzon, hogy a sokféle színt létrehozza.67
Sokféle ilyen genetikai kapcsoló létezhet, amik a variációt, illetve az adaptációt elősegítik. Ennek ellenére az evolucionisták nem tudják megmagyarázni a létezésüket. Az evolúció összes bizonyítéka nem más, mint a meglévő kapcsolók megváltozása.
A „funkciónövekedéssel járó mutáció” úgy hangzik, mintha egy újabb tulajdonságot hozna létre az élőlényben, nem pedig, hogy valamit összetörne. Az evolucionisták azonban a funkciónövekedéssel járó mutációt úgy határozzák meg, mint „egy olyan mutáció, ami új, vagy megnövekedett hatást kölcsönöz egy fehérjének”. Dr. Lightner több példát is említ.68 Az egyik a pajzsmirigy hormontermelését szabályozza.
Adott egy receptor, ami a hormon termelését szabályozza, de mutációt szenved, így bekapcsolva marad, és nem válaszol a pajzsmirigyet stimuláló hormonra, tehát a pajzsmirigy többlet hormont termel. „A fehérje fokozott aktivitása”? Igen ám, de a pajzsmirigy fokozott hormontermelése betegséget okoz.
Lightner azt írja, hogy:
a funkciónövekedés meghatározásának félrevezető meghatározása ellenére semmiféle információnövekedés vagy javulás nem jelenik meg a biokémiai utakban. Ilyen biokémiai utak kifejlesztésének mechanizmusa nélkül, az evolúció nem más, mint mítosz.
Az epigenetika a természetes szelekcióba belezavar
Az epigenetika felfedezése a legutóbbi időkben több problémát jelent a természetes szelekció számára.
Az epigenetika a génaktivitás olyan módon történő módosítását jelenti, amikor egy metil csoportot (-CH3) bizonyos nukleotidokra (citozinra) tesznek rá a DNS-ben. Ezt a folyamatot metilációnak nevezik. A terjedelmes metilcsoport a transzkripciós gépezetet (RNS polimeráz) gátolja, így egy metilált gén nem termel semmiféle fehérjét (vagyis, „le van némítva”).
A metilációt környezeti hatások váltják ki. Gyűlnek a bizonyítékok arra, hogy a metilált gének továbbadódnak generációról generációra.69 A metiláció megfordítható. Az epigenetikai öröklődés példái közé tartozik a sziromlevelek száma a növényekben illetve a szőr színe az egerekben. Egerekben bizonyos körülmények között az anyaegér táplálkozási szokásai az utódainak a szőrszínét befolyásolja. A szőrszín az unokákig képes öröklődni, de ez a hatás több generáció után megszűnik, ha a táplálkozás megváltozik. A környezeti hatás megszűnésével a metilációs mintázatok visszaállnak az eredeti állapotukba.70
Ilyen epigenetikai hatások a természetes szelekció magyarázatát még jobban megnehezítik, mivel mozgó céltáblát jelentenek. Dr. Robert Carter ezt így magyarázza:
Tegyük fel, hogy az egerek táplálkozási szokásai olyan epigenetikai változásokat hoznak létre az utódokban, hogy a szőrszínük jó álcázást tesz lehetővé. Azok az egerek, amelyek nem rendelkeznek ezzel a színnel, hátrányt szenvedhetnek. Ámbár semmiféle különbség nem rejlik azokban a génekben, amelyek a szőrszínt meghatározzák. Az egereknek attól más a színe, mert valamit ettek. Egyes egyedek úgy pusztulnának el, hogy nem is volt genetikai hátrányuk, és amint a hatás megszűnik, hiába volt az a sok szelekció.71
Az epigenetika így tehát több „zajt” kelt, mint amit a természetes szelekció „hall”. A sok láthatatlan, enyhén káros mutáció, illetve a kevés hasznos mutáció láthatatlansága miatt nem csoda, hogy kevés példát látunk a természetes szelekcióra.
A természetes szelekció túl lassú
A híres evolucionista genetikus J.B.S. Haldane (1892-1964) egyike volt annak a három embernek, aki megalkotta a populációgenetikát. Haldane súlyos problémát fedezett fel az evolúcióelméletben, és egy kulcsfontosságú cikkben 1957-ben le is írta, és „a szubsztitúció költségének” nevezte.72 Ha egy jótékony mutáció előfordul egy populációban, a gyakoriságának meg kell növekednie ahhoz, hogy a populáció fejlődjön (ha a mutáció pusztán egyetlen egyedben marad, akkor az evolúció nem tud haladni, ez nyilvánvaló). Más szavakkal: helyettesítenie kell a nem-mutált géneket a populációban. De ez csak korlátozott sebességen tud működni. Egy fontos tényező a helyettesítés sebességében a faj keletkezési rátája. Egy emberféle élőlénynek, ahol a generációs idő körülbelül 20 év, és ahol alacsony a szaporodási ráta, a mutációk számának növekedése egy populációban elég lassú.
Képzeljünk el 100.000 majmot, ami az ember feltételezett őse. Képzeljük el azt is, hogy egy hím és egy nőstény is olyan jótékony mutációban részesül, hogy ezzel mindenki mást túléltek, a többi 99.998 egyed pedig mind kihalt. És ezek után a fennmaradó párnak sikerült újra benépesíteni az egész populációt egyetlen generáció alatt! Ráadásul ez minden generációban megismétlődött (20 évenként), 10 millió éven keresztül, ami több, mint az ember és a csimpánz közös ősének feltételezett kora. Ez azt jelenti, hogy 500.000 (10 millió/20) jótékony mutációt adtak a populációhoz. Még ezzel a teljesen irreális elgondolással, ami az evolúció folyamatát maximalizálja, az emberi genomnak csakis a 0.02%-át lehet generálni. Ha figyelembe vesszük, hogy az ember és a csimpánz (mint állítólagos legközelebbi élő rokonunk) DNS-e között a különbség legalább 5%, vagyis 150 millió bázis73 („betű”), az evolúció elég nyilvánvaló probléma előtt áll, hogy az emberben lévő genetikai információ eredetét megmagyarázza. Innen származik a „hulladék DNS” elképzelés: amikor az evolucionisták felismerték, hogy a természetes szelekció nem tud elég DNS-t létrehozni, vagyis a nagy többség nem rendelkezik funkcióval (a következő fejezetben lesz még több szó a hulladék DNS-ről).

Ámbár ha realisztikusabb fitnessz/szelekciós illetve szaporodási rátát használunk, akkor a jótékony mutációk száma lecsökken. Haldane azt is kiszámolta, hogy nem több mint 1.667 jótékony mutáció mehetett végbe az állítólagos 10 millió év alatt, ami eltelt az ember és a majom közös őse óta. Ez pusztán egyetlen mutáció 300 generációnként átlagosan. Mindazt, ami egyedileg emberré tesz minket, meg kell magyarázni ezekkel a mutációkkal.
Abban az értelemben, ahogyan Haldane ezt a szót használta (a populációgenetikában) egy szubsztitúció egy olyan mutációs esemény, ami az egész populációban elterjed (vagyis „fixálódik”). Ez lehet génduplikáció vagy kromoszóma inverzió, vagy egyetlen nukleotid szubsztitúció, inszerció (beépülés), vagy deléció (törlés). A biológusok azt találták, hogy a szubsztitúciók túlnyomó többsége valóban egyetlen nukleotid szintjén történik, így Haldane számítása nagy nehézséget állít annak, hogy mi mehet végbe az evolúció során, mivel 1.667 nukelotid szubsztitúció összege nem hoz ki egy átlagos méretű gént.
Azt is meg kell jegyezni, hogy Haldane sok egyszerűsítő feltételezést állított fel, ami a szubsztitúciók lehetséges számát optimalizálja. Ez azt jelenti, hogy a lehetséges szubsztitúciók száma sokkal kisebb is lehet, mint amit Haldane kiszámolt.
Haldane analízise azt mutatja, hogy a mutációk természetes szelekciója nem tudja létrehozni az emberi genomot; ehhez túlságosan lassú. Az evolucionisták igyekeztek figyelmen kívül hagyni ezt a problémát, de nemrég Walter ReMine újra felelevenítette a problémát egy újabb számolással, és Haldane több félreértését is tisztázta.74
A természetes szelekció, ha egyáltalán működik, a teremtést támasztja alá
A természetes szelekció ettől még működik? Néhány esetben igen, mint ahogyan azt több példában láttuk. De egyszerűen túl sokat tulajdonítanak neki. A természetes szelekció néhány klasszikus példája, mint a páva farka, amiről Darwin azt feltételezte, hogy azért jött létre, mert a nőstények a hímek látványos farkát kedvelték („szexuális szelekció”), az nem más, mint kitalált mese.75

Még akkor is, ha a szelekció a kísérletes igazolást is kiállta volna, nem tudja megmagyarázni például a farok eredetét76, legfeljebb annak a karbantartását, hiszen a természetes szelekció egy fenntartó erő, ahogyan azt eredetileg a 19. századi teremtéshívő Blyth feltételezte.
Ez mit jelent?
Azt, hogy a mutációk, illetve a természetes szelekció elégtelen mechanizmus az evolúció általános elméletére. A molekuláris biológiában történő robbanás az evolúció, mint a földön lévő diverzitás magyarázata elleni bizonyítékokat csak megnövelte. A következő fejezetben megvizsgáljuk, ami az evolúcióelmélet mögött van, vagyis a genetika területét, és hogy a genetikában történt legújabb felfedezések hogyan ingatják meg az evolúció általános elméletét.
Bővebben a könyvről: Evolution’s Achilles’ Heels
Hivatkozások:
1. Darwin, C.R. Az állatok és növények sokfélesége a háziasítás során, 1. kiadás, 1. kötet, 1. nyomtatás, John Murray, London, UK, 6. o., 1868.
2. Darwin ilyen pintyeket figyelt meg a Galápagos szigeteken. Lásd Wieland, C., Darwin’s finches, Creation 14(3):22–23, 1992; creation.com/darwins-finches.
3. Kerkut, G.A., Implications of Evolution, Pergamon, Oxford, UK, p. 157, 1960.
4. Lewin, R., Evolutionary theory under fire, Science 210(4472):883–887, 1980.
5. Lewin, Evolutionary theory under fire, p. 884.
6. Endler, J. A., Natural Selection in the Wild, Princeton University Press, NJ, USA, p. 5, 1986.
7. Dawkins, R., The Greatest Show on Earth, Free Press, New York, p. 139, 2009. Szintén lásd Sarfati, J., Dawkins playing bait and switch with guppy selection, February 2010; creation.com/dawkins-guppy.
8. Lásd Catchpoole, D., Defining terms, January 2011; creation.com/defining-terms.
9. Endler, Natural Selection in the Wild, p. 8.
10. Endler, Natural Selection in the Wild, p. 245.
11. Endler, Natural Selection in the Wild, p. 246.
12. A szexuális szelekció az a természetes szelekció olyan változata, amikor egy fajban a hím vagy a nőstény preferenciát mutat egy olyan partner felé, aki bizonyos tulajdonságokat mutat (mint például a testszín).
13. Waggoner, B., Carl Linnaeus (1707–1778), 2000-ben felújítva; ucmp.berkeley.edu.
14. Paul Pearson áttekintése a Nature 425(6959):665, 2003 számában. Pearson azt mondta Huttonról, hogy „a szelekció mechanizmusát arra használta, hogy a természetben lévő összes változatosságot megmagyarázza”, noha „különösen elvetette az evolúciót, mint „romantikus fantáziát”.”
15. Pearson, Nature 425(6959):665, 2003.
16. Gould, S.J. idézete, A természetes szelekció mint alkotó erő, Az evolúciós elmélet szerkezete, Belknap Press, Harvard University, Cambridge, MA, USA, 138. o., 2002.
17. Gould, S.J. idézete, A természetes szelekció, mint alkotó erő.
18. Pearson, Nature 425(6959):665, 2003.
19. Uo.
20. Eiseley, L., Darwin és a titokzatos X úr, E.P. Dutton, New York, NY, USA, p. 55, 1979.
21. Grigg, R., Darwin törvénytelen gyermeke, Creation 26(2):39–41, 2004; creation.com/brainchild.
22. Blyth, E., Eiseley művében idézve, Darwin és a titokzatos X úr, 108. o.
23. Bergman, J., Evolutionary naturalism: an ancient idea, J. Creation 15(2):77–80, 2001; creation.com/naturalism-old.
24. Bergman, J., Did Darwin plagiarize his evolution theory? J. Creation 16(3):58–63, 2002; creation.com/darwin-plagiarize-evolution.
25. Waggoner, Carl Linnaeus (1707–1778).
26. Például azok az egyházi személyek, akik a fiatal Darwint a Cambridge-i egyetemen oktatták: lásd Grigg, R., Darwin mentorai, Creation 32(1):50–52, 2009; creation.com/clergy-mentor-darwin.
27. Wieland, C., könyv áttekintés: The Beak of the Finch; Evolution in Real Time, J. Creation 9(1):21–24, 1995; creation.com/beak_finch.
28. A mezozoikum korabeli kövekről egy katalógus („a dinoszauruszok kora”), lásd: Werner, C., Evolúció: a nagy Kísérlet, 2. kötet – az élő kövületek, New Leaf Press, Green Forest, AR, USA, 2009.
29. Emiatt a brit paleontológus Simon Conway Morris szerint „egyszer mindnyájan férgek voltunk” (Új Tudós szaklap) 179(2406):34, 2003.
30. Marsh, F.L., A természetbeli variáció és állandóság, Pacific Press, Mountain View, CA, USA, p. 37, 1976.
31. Scherer, S., Basic Types of Life, in Scherer, S. (ed.), Typen des Lebens, Pascal-Verlag, Berlin, 197. o., 1993; Dembski, W.A., Mere Creation, InterVarsity Press, Downers Grove, IL, USA, 8. fej., 1998.
32. Batten, D., Ligerek és wholphin-ok? Mi van itt? Creation 22(3):28–33, 2000; creation.com/liger.
33. Frair, W., Kreacionista hallgatóként ilyen irányú tapasztalataim voltak több egyetem állatrendszertani tanszéken 2011; márciusában; creation.com/frair.
34. Lightner, J.K., Identification of species within the sheep-goat kind (Tsoan monobaramin), J. Creation 20(3):61–65, 2006; creation.com/tsoan.
35. Stevison, L.S., Hoehn, K.B., and Noor, M.A.F., Effects of inversions on within-and between-species recombination and divergence, Genome Biol. Evol. 3:830 –841, 2011.
36. Catchpoole, D. and Wieland, C., Speedy species surprise, Creation 23(2):13–15, 2001; creation.com/speedy-species-surprise.
37. Reznick, D.N. et al., Evaluation of the rate of evolution in natural populations of guppies (Poecilia reticulata), Science 275(5308):1934–1937, 1997.
38. Morell, V., Predator-free guppies take an evolutionary leap forward, Science 275(5308):1880, 1997.
39. Uo.
40. Arthur Jones Biology, in Ashton, J. (ed.), In Six Days—Why 50 Scientists Choose to Believe in Creation, Master Books, Green Forest, AR, USA, 2001, pp. 241–248; creation.com/jones. Erre szép ábrázolása van az afrikai tavakban élő cichlid halakra a következő publikációban: Kocher, T.D. et al., Similar morphologies of cichlid fish in Lakes Tanganyika and Malawi are due to convergence, Molecular Phylogenetics and Evolution 2(2):158–165, 1993. A következő publikációban szintén megtekinthető: Williams, A., Taking firm hold of an illusion, J. Creation 20(3):45–49, 2006; creation.com/Falk.
41. Edwards, C.J. et al., Ancient hybridization and an Irish origin for the modern polar bear matriline, Current Biology 21:1251–1258, 2011.
42. Sarfati, J., Design in living organisms (motors: ATP synthase), J. Creation 12(1):3–5, 1998; creation.com/motor and Thomas, B., ATP synthase, Creation 31(4):21–23, 2009; creation.com/atp-synthase.
43. Lásd description and animation in Smith, C., Fantastic voyage, Creation 30(1):20–23, 2007; creation.com/fantastic-voyage.
44. Christen, B. et al., The essential genome of a bacterium, Molecular Systems Biology 7:528, 2011. A Caulobacter crescentus genomjához szükséges részek 8 bp-os felbontásnál 480 ORF-ot (open reading frame, vagyis nyitott olvasási keret, ami fehérjét kódol) eredményezett, 402 szabályozó szekvencia és 130 nemkódoló elem, magába értve 90 intergenikus szegmens, aminek ismeretlen a funkciója; 492,941 bp összességében. Érdekes módon, abból a 320 létszükséges fehérjéből, ami mind az Caulobacter és az E. coli genomban megtalálható, ebből több mint a harmada nem szükséges az E. coli-ban. Ettől függetlenül, az E. coli a saját létszükséges fehérjéivel rendelkezik, amelyek nincsenek jelen a Caulobacter genomban, illetve nem szükségesek a Caulobacter számára. A szerzők azt a következtetést vonták le, hogy egy baktérium létszükséges genomja az összes komponensének összjátékából áll össze a baktériumok esetében. Így például az ATP szintáz szükséges a Caulobacter számára, de nem az E. coli számára, mivel az utóbbi a fermentáció révén tudja az ATP-t létrehozni.
45. Az IAEA és a FAO évente kétszer a Növényi Mutációs Riportokat közösen is publikálják.
46. Pl. Catchpoole, D., Morning glory’s designer label clothing, Creation 29(1):49–51, 2006; creation.com/morning-glorys-designer-label-clothing.
47. Mira A., Ochman H., and Moran N.A., Deletional bias and the evolution of bacterial genomes, Trends in Genetics 17(10):589–596, 2001.
48. Batten, D., Bacteria „evolving in the lab”? June 2008; creation.com/lenski.
49. Blount, Z.D. et al., Genomic analysis of a key innovation in an experimental Escherichia coli population, Nature 489:513–518, 2012.
50. See review Batten, D., Clarity and confusion – A review of The Edge of Evolution, J. Creation 22(1): 28–33, 2008; creation.com/edge-evolution. Behe a mutáns citrát fogyaztó E. coliról is írt kommentárt. Behe, M., Rose-colored glasses: Lenski, citrate, and BioLogos, 13 Nov 2012; www.evolutionnews.org/2012/11/rose-colored_gl066361.html.
51. omim.org
52. Haldane, J.B.S., The cost of natural selection, J. Genetics 55:511–524, 1957; Maynard Smith, J., The Theory of Evolution, Penguin Books, Harmondsworth, UK, p. 239, 1958.
53. Sanford, J. Batten, D. növénygenetikus interjújában: „Darwinian evolution is impossible”, „Darwini evolúció lehetetlen”, Creation 30(4):45–47, 2008; creation.com/sanford.
54. Sanford ezt az analízist a Genetika Entrópia és a Genom Rejtélyei c. könyvében mutatta be
FMS Publications; 3. kiadás, 2008 márciusa (a creation.com honlapon megtalálható). Az első kiadás áttekintése, itt elolvasható: Truman, R., From ape to man via genetic meltdown: a theory in crisis – A review of Genetic Entropy & The Mystery of the Genome by John C. Sanford, J. Creation 21(1):43–47, 2007; creation.com/sanford-áttekintés.
55. See Sanford, J., Critic ignores reality of Genetic Entropy, March 2013; creation.com/genetic-entropy.
56. Mendel’s Accountant; (Mendel Könyvelője) mendelsaccountant.info; Sanford, J. et al., Mendel’s Accountant, SCPE 8(2): 147–165, 2007; www.scpe.org/index.php/scpe/article/view/407/77.
57. Wieland, C., Blind fish, island immigrants and hairy babies, Creation 23(1):46–49, 2000; creation.com/blind-island.
58. Sarfati, J., Christopher Hitchens-blind to salamander reality, July 2008; creation.com/hitchens.
59. Wieland, C., Beetle bloopers, Creation 19(3):30, 2003; creation.com/beetle-bloopers.
60. Wieland, C., Superbugs not super after all, Creation 20(1):10–13, 1997; creation.com/superbugs.
61. Wieland, C., Rapid tomcod „evolution by pollution”? February 2011; creation.com/tomcod.
62. Catchpoole, D., The Stickleback: Evidence of evolution? (A tüskés pikó: Az evolúció bizonyítéka?) September 2009; creation.com/stickleback.
63. Pennisi, E., Evolutionary biology: Changing a fish’s bony armor in the wink of a gene, Science 304(5678):1736–1739, 2004.
64. Coyne, J.A., Switching on evolution – how does evo-devo explain the huge diversity of life on Earth? Nature 435(7046):1029–1030, 2005.
65. Carroll, Catchpoole művében idézve.
66. Catchpoole, D., A tüskés pikó
67. Lightner, J., interviewed by Batten, D., Getting it right, Creation 32(3):40–43, 2010; creation.com/creationist-veterinarian.
68. Lightner, J., Gain-of-function mutations, J. Creation 19(3):7–8, 2005; creation.com/gain-of-function.
69. Lásd pl., Whitelaw, E., Sins of the fathers, and their fathers, European J. Human Genetics 14:131–132, 2006; Pembrey, M.E. et al., Sex-specific, male-line transgenerational responses in humans, European J. Human Genetics 14:159–166, 2006; Jimenez-Chillaron, J.C. et al., Intergenerational transmission of glucose intolerance and obesity by in utero undernutrition in mice, Diabetes 58:460–468, 2009.
70. Morgan, H. et al., Epigenetic inheritance at the agouti locus in the mouse, Nature Genetics 23:314–318, 1999.
71. Carter, R., Darwin’s Lamarckism vindicated? March 2011; creation.com/epigenetics-and-darwin.
72. Haldane, J.B.S., The cost of natural selection, J. Genetics 55:511–524, 1957.
73. See Tomkins, J. and Bergman, J., Genomic monkey business-estimates of nearly identical human-chimp DNA similarity re-evaluated using omitted data, J. Creation 26(1):94–100, 2012; creation.com/chimp. The difference may even be as high as 30%, as geneticist Richard Buggs showed: Chimpanzee? October 2008; www.refdag.nl/chimpanzee_1_282611.
74. Batten, D., Haldane’s dilemma has not been solved, J. Creation 19(1):20–21, 2005; creation.com/haldane. See also ReMine, W., Cost theory and the cost of substitution-a clarification, J. Creation 19(1):113–125, 2005; creation.com/cost.
75. Catchpoole, D., Peacock tail tale failure, June 2008; creation.com/peacock-tail-tale-failure.
76. Burgess, S., A páva farok tollának szépsége és a nemi szelekció elmélet problémái, J. Creation 15(2):94–102, 2001; creation.com/peacock.